/home/virtual/kjpt/journal//../xmls/kjpt-45-4-380.xml
한국산 마속(마과)의 외부형태형질에 의한 분류학적 연구
적 요
한국산 마속(Dioscorea L.) 6분류군(마; D. polystachya, 참마; D. japonica, 부채마; D. nipponica, 단풍마; D. quinqueloba, 각시마; D. tenuipes, 도꼬로마; D. dokoro)을 대상으로 외부형태학적 형질을 관찰하였다. 지하부의 유형, 주아의 형성 유무 등이 마절(Enanatiophyllum)과 부채마절(Stenophora)을 구분하는 유용한 형질로 관찰되었으며, 엽병 기부의 돌기 유무, 꽃의 형태 및 색, 열매의 형태, 종자 날개의 형태 등을 비롯한 약의 위치 와 암술 형태와 같은 미세구조적 형질 등이 종을 식별하는 중요한 형질로 판단되었다. 또한 상기 식별형질을 바탕으로 검색표를 제시하였고, 한반도 내 마속 식물의 분포를 확인하였다.
주요어: 마속, 마과, 외부형태, 검색표, 분포
Abstract
The morphological characters were observed in six taxa in the genus Dioscorea of Korea (D. polystachya, D. japonica, D. nipponica, D. quinqueloba, D. tenuipes, and D. dokoro). The underground structure and the formation of bulbil were useful characters for separating sect. Enanatiophyllum from sect. Stenophora. The important characters for discriminating taxonomic entities were as follows: the position of the anther and the shape of the pistil in the microstructural characters, including the protuberance of the base of the petiole, the shape and color of the flower, the shape of the fruit, and the shape of the seed wing. Based on the results, a key to Korean Dioscorea is provided and the distribution of these species in Korea is described.
Keywords: Dioscorea, Dioscoreaceae, morphology, key, distribution
마속( Dioscorea L.)은 백합목(Liliales), 마과(Dioscoreaceae)에 포함되는 분류군으로 주로 열대지역, 동아시아, 지중해 연안 및 남·북아메리카 등에 약 600종이 분포한다( Coursey, 1969; Li et al., 1978; Ohwi, 1984; Im, 2000; Lee, 2006). 마속은 Linnaeus (1753)에 의해 잎의 형태적 차이로 처음 기재된 후 분류학적 한계설정에 많은 이견과 혼동이 있어 왔다. Burkill (1960)은 마과를 자웅동주와 자웅이주의 형질로 Paradioscreeae와 Dioscoreeae의 2족으로 나누고, 지하부의 유형, 열매 및 종자의 형태를 바탕으로 마속을 23개 절로 구분하였다. 또한 Beccari (1870)는 자방의 1실속에 포함되어 있는 종자를 바탕으로 마속을 2개의 종자가 포함된 마속과 그 이상인 Stenomeris속( Stenomeris Planch.)으로 구분하였으며, Knuth (1850)는 종자에 발달되는 날개의 특징으로 종자를 원형으로 둘러싼 형태인 Eudioscorea아속( Eudioscorea Kunth)과 한쪽으로 발달하는 Helmia아속( Helmia Kunth)으로 구분하였다. 그리고 Engler and Prantl (1897)는 지하부의 유형, 잎의 형태, 꽃의 형태 및 종자의 형태 등의 형질을 바탕으로 39개 절로 구분하였으며, 이의 분류체계는 현재 널리 적용되고 있다. 이외에도 Matsuno (1918), Maekawa et al. (1962) 등의 다수 학자들은 잎의 형태, 잎차례, 주아의 유무, 엽병 기부형태, 꽃의 형태 및 색, 종자의 형태 등을 속 내 분류체계를 결정하는 주요 형질로 취급하였으며, 최근 지하부와 열매의 색소체 유전자를 이용한 계통분석( Wilkin et al., 2005) 및 DNA염기서열을 이용한 분자생물학적 연구가 이루어지면서 분류군간의 유연관계와 진화양상이 논의되고있다( Gao et al., 2008; Hsu et al., 2013; Gawande et al., 2015). 한국산 마속에 대한 연구는 Palibin (1901)의 각시마( D. tenuipes Franch. & Sav.)에 대한 기록을 시작으로 Nakai (1911, 1952)는 7종, Mori (1921)는 8종 1변종, Park (1949)은 7종 1변종을 기록하고 있으며, 현재는 마( D. batatas Decne.), 참마( D. japonica Thunb.), 부채마( D. nipponica Makino.), 단풍마( D. quinqueloba Thunb.), 각시마( D. tenuipes Frnach. & Sav.), 도꼬로마( D. tokoro Makino.), 국화마( D. septemloba Thunb.), 둥근마( D. bulbifera L.) 등 8종이 기록되고 있다( Lee, 2003; Lee, 2006; Oh and Lee, 2007). 하지만 이는 단순한 학명의 기록 또는 도감의 기재로서 마속 분류군의 형태적 유사성 및 개체 내 심한 변이의 폭으로 인하여 분류학적 처리에 어려움이 있는 실정이다. 최근 Kang et al. (1992)과 Oh et al. (1995, 1996)에 의해 한국산 마속의 분류학적 연구가 진행되었으나, Kang et al. (1992)은 참마의 재료가 생략된 상황에서 문헌상의 식별 형질로만 참마를 구분하여 마와 참마를 혼동한 채 연구를 수행하였고, Oh et al. (1995, 1996)의 연구는 분류군간의 뚜렷한 식별형질을 제시하지 못하였으며, 특히 현재 국내에서 마의 학명을 D. batatas ( Decaisne, 1854)와 D. polystachya ( Turczaninow, 1837)를 혼용하여 사용하는 등 최근의 분류학적 연구결과를 반영하지 못하고 있는 실정이다. 따라서 본 연구의 목적은 한국산 마속의 전반적인 외부형태 및 미세구조의 형질을 재검토하고, 분류군간의 유용한 식별 형질을 파악하여 종의 명확한 한계를 구분하는 데 있다.
재료 및 방법
재료
실험재료는 2010년 5월부터 2015년 10월까지 개화기와 결실기를 중심으로 채집된 성숙한 개체를 FAA (Formalinacetic acid-ethyl alcohol)에 고정한 액침표본과 안동대학교 생약자원학과(ANH), 국립수목원(KH), 서울대학교 산림과학부(SNUA)에 소장되어 있는 건조표본을 사용하였다. 또한 일부는 안동대학교 실험포장에 이식하여 재배한 재료를 사용하였으며, 관찰 및 실험과 분포 확인을 위해 사용된 표본은 Appendix 1에 기재하였다. 또한 현재까지 관찰 및 채집이 이루어지지 못한 국화마(D. septemloba) 및 둥근마(D. bulbifera)는 본 연구에서 제외하였다.
방법
동일집단에서 성숙개체를 대상으로 건조표본, 액침표본, 생체를 이용하여 영양기관 및 생식기관의 정성·정량적 형질을 관찰(해부현미경; Olympus Ax-70, Japan; Olympus DP2-BSW, Germany), 측정 및 촬영하였고, 주요형질을 도해하였다. 미세구조 관찰을 위하여 FAA에 고정되어 있는 성숙개체의 암술과 수술을 취하여 1.5% glutaraldehyde에 전처리 후 0.1 M phosphate buffer (pH 6.8)로 2회 세척, 30−100% ethanol의 단계적인 탈수과정을 거처 isoamylacetate에 2회 치환하였다. 이를 critical point dryer (HCP-2)로 건조 후 ion sputter로 gold coating (thickness: 200−250 A)한 후 주사전자현미경(Hitach; S-2500C, Japan; 20 Kv; 80 W; working distance 15 mm)으로 관찰 및 촬영하였다.
결 과
1. 지하부 구조
마절( Enanatiophyllum)의 마와 참마는 수직으로 곧게 뻗는 괴근의 형태로 발달하며, 부채마절( Stenophora)의 부채마, 단풍마, 각시마, 도꼬로마는 수평으로 퍼지는 근경의 형태로 발달한다( Fig. 1, Table 1).
2. 주아
마절의 마와 참마는 엽액에 주아가 형성되고, 부채마절의 부채마, 단풍마, 각시마, 도꼬로마는 주아가 형성되지 않았다( Fig. 2, Table 1).
3. 잎
1) 엽병
엽병의 길이는 도꼬로마가 평균 6.7 cm로 가장 길고, 각시마가 평균 3.1 cm로 가장 짧게 관찰되었다. 또한 단풍마, 각시마의 엽병 기부에 한 쌍의 작은 다육성 돌기가 관찰된 반면 마, 참마, 부채마와 도꼬로마는 관찰되지 않았다( Fig. 3, Table 1).
2) 잎의 형태
잎의 길이는 단풍마가 평균 9.7 cm로 가장 길고, 마는 평균 5.6 cm로 가장 짧게 관찰되었고, 너비는 단풍마가 평균 8.7 cm로 가장 넓고, 각시마는 평균 4.4 cm로 가장 좁게 관찰되었다. 또한 마는 삼각형, 참마는 비율이 2:1 미만인 좁은 삼각형, 부채마와 단풍마는 삼각상 난형, 각시마는 삼각상 심장형, 도꼬로마는 심장형으로 관찰되었으며, 마와 참마는 0−1쌍의 측열편이 갈라지고, 부채마와 단풍마는 1−4쌍의 측열편이 깊게 갈라지며, 각시마와 도꼬로마는 갈라짐이 관찰되지 않았다( Fig. 4, Table 1).
3) 부채마와 단풍마 잎의 비교
부채마와 단풍마의 잎을 중앙열편(middle-lobe), 1측열편(first-lobe), 2측열편(second- lobe)으로 구분하여 비교하였다( Fig. 5). 중앙열편의 선단의 형태는 부채마와 단풍마 모두 예두로 관찰되었고, 1측열편과 2측열편의 선단의 형태는 부채마가 예두, 단풍마는 둔두로 관찰되었다. 중앙열편의 길이는 부채마가 평균 5.4 cm, 단풍마는 평균 5.3 cm, 너비는 부채마가 평균 3.3 cm, 단풍마는 평균 3.9 cm로 관찰되었다. 1측열편의 길이는 부채마가 평균 2.1 cm, 단풍마는 평균 2.0 cm, 너비는 부채마가 평균 1.4 cm, 단풍마는 평균 2.0 cm로 관찰되었다. 2측열편의 길이는 부채마가 평균 1.6 cm, 단풍마는 평균 1.3 cm, 너비는 부채마가 평균 1.4 cm, 단풍마는 평균 1.7 cm로 관찰되었다. 또한 중앙열편과 1측열편 사이의 각은 부채마가 평균 57º, 단풍마는 평균 63º로 관찰되었고, 1측열편과 2측열편 사이의 각은 부채마가 평균 77o , 단풍마는 평균 70º로 관찰되었다( Fig. 5, Table 2).
4. 꽃
단성화이며, 3장씩의 내·외화피로 구성된다. 개화하는 시기는 5−9월이다.
1) 화서
마와 참마는 자·웅화 모두 수상화서의 형태로 관찰되었으나, 마의 자화경은 지그재그 형태로 발달하였고, 참마는 곧게 발달하였다. 부채마, 단풍마, 각시마와 도꼬로마는 자·웅화 모두 총상화서로 관찰되었다( Fig. 6, Table 2).
2) 화피의 형태
개화시 화피가 펼쳐지지 않는 마와 참마, 종형으로 개화하는 부채마, 단풍마 및 도꼬로마, 화피가 뒤로 젖혀지는 각시마로 관찰되었으며, 마와 참마는 화피편의 형태가 넓은 난형, 도꼬로마는 난형, 부채마와 단풍마는 도란형, 각시마는 선형으로 관찰되었다( Figs. 7− 8, Table 1).
3) 화피의 색
마와 참마는 꽃의 색이 모두 흰색으로 관찰되었으며, 부채마는 옅은 황색, 단풍마는 짙은 황색, 각시마는 연녹색, 도꼬로마는 상아색으로 관찰되었다( Table 1).
4) 약의 위치
마와 참마 및 부채마는 약이 수술의 전면, 단풍마, 도꼬로마는 약이 수술의 측면, 각시마는 수술의 정단에 위치하는 것으로 관찰되었다( Fig. 9).
5) 암술의 형태
각시마는 암술대의 기부에서 3갈래로 갈라져 분지하였고, 다른 분류군은 암술대의 정단부에서 3갈래로 분지하는 것으로 관찰되었다( Fig. 9).
5. 열매 및 종자
1) 열매
열매의 길이는 도꼬로마가 평균 23.5 mm로 가장 길고, 단풍마는 평균 13.1 mm로 가장 짧게 관찰되었고, 너비는 도꼬로마가 평균 27.6 mm로 가장 넓고, 부채마는 평균 15.9 mm로 가장 좁게 관찰되었다. 또한 열매의 형태는 마는 도란상구형, 부채마는 장타원형, 참마 및 단풍마, 각시마, 도꼬로마는 넓은 도란형의 형태로 관찰되었다( Fig. 10, Table 1).
2) 종자
종자의 길이는 부채마가 평균 5.6 mm로 가장 길고, 마는 평균 3.9 mm로 가장 짧게 관찰되었고, 너비는 참마가 평균 4.2 mm로 가장 넓고, 부채마는 평균 3.4 mm로 가장 좁게 관찰되었다. 종자는 주변에 막질의 날개를 가지고 있으며, 날개의 길이는 부채마가 평균 13.6 mm로 가장 길고, 단풍마는 평균 8.2 mm로 가장 짧게 관찰되었다. 너비는 참마가 평균 15.8 mm로 가장 넓고, 부채마는 평균 6.6 mm로 가장 좁게 관찰되었다. 또한 마와 도꼬로마는 반구형(domed), 부채마는 장타원형, 참마 및 단풍마, 각시마는 구형의 형태로 관찰되었으며, 종자가 막질날개의 기부에 위치하는 부채마와 중심부에 위치하는 그 외 분류군으로 관찰되었다( Fig. 10, Table 1). 이상의 결과를 바탕으로 작성한 한국산 마속의 검색표는 다음과 같다.
한국산 마속의 검색표
1. 지하부는 수직으로 뻗는 괴근이며, 주아가 형성되고, 수상화서이다.
2. 잎은 삼각형, 자화경은 지그재그형태로 발달하며, 열매의 너비는 16.6−17.9 mm이다 ···· 마 D. polystachya
2. 잎은 좁은 삼각형, 자화경은 곧게 발달하며, 열매의 너비는 24.6−29.5 mm이다 ··············· 참마 D. japonica
1. 지하부는 수평으로 뻗는 근경이며, 주아가 형성되지 않고, 총상화서이다.
3. 잎은 1−4쌍의 측열편이 갈라진다.
4. 엽병의 기부에는 다육성 돌기가 발달하고, 잎의 측열편은 둔두, 종자는 막질 날개의 중앙에 위치한다 ····································· 단풍마 D. quinqueloba
4. 엽병의 기부에는 다육성 돌기가 발달하지 않고, 잎의 측열편은 예두, 종자는 막질 날개의 기부에 위치한다 ································· 부채마 D. nipponica
3. 잎은 측열편이 갈라지지 않는다.
5. 엽병의 기부에는 다육성 돌기가 발달하고, 잎은 삼각상 심장형, 화피는 뒤로 젖혀지며, 약은 수술대의 정단에 위치하고, 암술은 기부에서 갈라진다 ·········································· 각시마 D. tenuipes
5. 엽병의 기부에는 다육성 돌기가 발달하지 않고, 잎은 심장형, 화피는 뒤로 젖혀지며, 약은 수술대의 측면에 위치하고, 암술은 정단에서 갈라진다 ······················································· 도꼬로마 D. tokoro
고 찰
한국산 마속(Dioscorea L.) 분류군에 대한 외부형태적 형질인 지하부의 유형, 주아, 잎의 형태 및 잎차례, 엽병 기부의 돌기 유무, 꽃의 형태 및 색, 암·수술의 형태, 열매의 형태, 종자 및 종자 날개의 형태 등을 비롯한 미세구조인 수술과 암술의 형태를 본 연구에서 재검토 하였다.
일반적으로 마속 내 절을 분류하는 형질 중 지하부의 유형( Engler and Prantl, 1897; Burkill, 1960)과 주아의 유무( Matsuno, 1918; Maekawa, 1962)는 수직으로 곧게 뻗는 괴근(tuber)의 형태를 가지고, 주아가 형성되는 마절(Enantiophyllum)과 수평으로 뻗는 근경(rhizome)의 형태를 가지고, 주아가 형성되지 않는 부채마절( Stenophora)로 관찰되어 절을 분류하는 형질로서의 가치가 있으나, 그 이외의 외부형태적 형질인 꽃의 형태, 열매 및 종자의 형태 등( Engler and Prantl, 1897; Burkill, 1960)은 한국산 마속에서는 절간의 뚜렷한 식별형질을 확인할 수 없었다. 마절( Enanatiophyllum)에 속하는 마( D. polystachya)와 참마( D. japonica)는 매우 유사한 분류군이며, 잎의 형태와 잎차례, 줄기 및 엽병의 색 등이 두 분류군을 식별하는 형질로 알려져 있다( Matsuno, 1918; Ohwi, 1984). 하지만 Kang et al. (1992)은 열악한 생육환경에서 자생하는 마와 참마는 식별형질인 잎의 형태나 잎차례에 차이가 없었고, 마의 줄기와 엽병은 색을 띄지 않아 종을 식별하는 형질로 생각하기에는 어려움을 있다고 보고하였으며, Oh et al.(1995)는 마와 참마는 같은 집단 및 동일 개체 내에서도 잎의 변이가 다양하여 잎의 형태로 마와 참마를 구별하는 것이 적합하지 않다고 보고한 바 있다. 마와 참마는 선행되어진 연구에서의 형질인 잎차례나 줄기 및 엽병의 색 등( Matsuno, 1918; Ohwi, 1984)에서 종간의 식별형질이 관찰되지 않았고, 그 외의 꽃의 형태, 내·외화피의 형태 및 크기, 종자의 형태 및 크기 등의 형질에서도 미세한 양적 형질의 차이만이 관찰되었을 뿐 종을 식별하는 분류형질로서 타당성을 확인하지 못했다. 하지만 잎의 형태가 마는 삼각형(deltate), 참마는 길이와 너비의 비율이 2:1 이상인 좁은 삼각형(narrowly-triangular)으로 확인되었고, 열매의 너비가 마는 16.6−17.9 mm, 참마는 24.6−29.5 mm로 확인되어 잎의 형태로 두 종을 분류한 Maekawa (1962)와 열매의 크기로 두 종을 분류한 Tominari (1984)의 언급과도 일치하였다. 또한 화경의 형태가 마의 자화경이 지그재그로 발달하여, 곧게 발달하는 참마와 차이점을 확인하였다. 부채마절( Stenophora)에 속하는 부채마( D. nipponica), 단풍마( D. quinqueloba), 각시마( D. tenuipes), 도꼬로마( D. dokoro)는 잎의 형태, 엽병 기부의 돌기 유무, 꽃의 형태 및 색, 열매 및 종자 날개의 모양 등의 외부형태에 의해 분류되고 있으며( Lee, 2003; Lee, 2006), 잎의 측열편이 깊게 갈라지는 부채마와 단풍마, 측열편이 갈라지지 않는 각시마와 도꼬로마로 구분된다( Oh et al., 1995). 잎의 측열편이 갈라지는 부채마와 단풍마는 잎의 형태적 유사성과 큰 폭의 변이로 인해 종의 식별에 어려움이 있다. 이에 부채마와 단풍마의 잎을 각 열편으로 구분지어 관찰하였으며, 각 열편의 길이와 너비, 열편 사이 각 등의 양적형질에 대한 차이는 미세하였으나, 측열편 선단의 형태가 부채마는 예두인데 반해 단풍마는 둔두의 형태를 띠어 두 종간의 차이점을 확인하였다. 또한 단풍마는 엽병의 기부에 돌기가 발달하여 그렇지 않은 부채마와 차이점을 확인하였으며, 꽃의 색, 종자의 형태 등의 형질에서 두 분류군간의 차이점을 확인하였다. 잎의 측열편이 갈라지지 않는 각시마와 도꼬로마는 잎의 형태가 삼각상 심장형인 각시마와 심장형인 도꼬로마로 구분되었으며, 각시마는 단풍마와 같이 엽병 기부에 돌기가 발달하여 그렇지 않은 도꼬로마와 차이점을 확인하였다. 또한 개화시 화피의 형태는 각시마는 선형이며 뒤로 젖혀지고, 도꼬로마는 난형이며 종형으로 개화하는 차이점을 확인하였다.
미세구조인 암술과 수술의 형태는 마절의 마와 참마 두 분류군 모두 암술대의 정단에서 3갈래로 분지하고, 약이 수술대의 전면에 위치하는 등 두 종간의 형태적 차이점은 확인되지 않았다. 반면 부채마절의 분류군에서는 암술과 수술의 다양한 형태를 확인하였다. 각시마는 암술대 정단에서 3갈래로 분지하는 다른 분류군 반해 기부에서 3갈래로 분지하였다. 또한 약이 수술대의 정면에 위치하는 부채마, 측면에 위치하는 단풍마와 도꼬로마, 상단부에 위치하는 각시마로 관찰되어 종을 분류하는 유용한 형질이라 판단된다.
현재 국내와 일본에서는 마의 학명을 Decaisne (1854)가 발표한 D. batatas를 사용하고 있는 반면 중국에서는 Turczaninow (1837)가 발표한 D. polystachya를 사용하고 있다. 두 학명은 지하부 유형, 주아의 존재, 잎의 형태 및 열매의 형태 등의 외부형태적 형질을 바탕으로 발표된 합법명이나, 선취권이 앞선 D. polystachya Turcz를 사용하는 중국의 학명처리가 타당한 것으로 판단된다. 상기 식별형질을 바탕으로 한반도 내 마속의 분포를 확인하였다. 마는 한반도 전역에 고루 분포하는 반면, 참마는 제주도 및 남해안 일대 섬 지역에서 한정적으로 분포하며, 부채마와 도꼬로마는 강원, 경기, 충북, 충남, 경북 등의 중부지역, 단풍마와 각시마는 전남, 경남, 제주도 등의 남부지역에 분포하는 것으로 확인되었다( Fig. 11). 단풍마와 참마는 한반도 전역에 분포한다고 알려져 있으나( Park, 1949; Lee, 2003; Lee, 2006; Oh and Lee, 2007), 이는 참마의 실체를 배재한 체 연구가 수행되어졌거나( Park, 1949) 마속의 각 분류군에 대한 식별형질의 부재( Lee, 2003; Lee, 2006; Oh and Lee, 2007)로 인해 정확한 동정이 이루어지지 않았기 때문이라 판단된다. 또한 국화마( D. septemloba)는 한반도의 남해안 지역과 일본의 Sinkoku, Kyushu지역에, 둥근마( D. bulbifera)는 한반도 전역을 비롯한 중국과 일본의 Honshu, Sinkoku, Kyushu 지역의 분포가 보고되고 있다( Ohwi, 1984; Ding and Gilbert, 2000). 그러나 국화마는 잎의 측열편이 깊게 갈라지며, 열매가 도란상구형인 특징, 둥근마는 잎이 심장형, 화피편의 형태가 피침형, 열매가 장타원형인 특징으로 동속 분류군과 구분되어지며, 본 연구를 통하여 ANU, KH, SNUA 등에 소장되어진 국화마와 둥근마로 동정된 표본을 확인한 결과, 잎의 변이로 인해 측열편이 깊게 갈라진 부채마와 단풍마가 국화마로 오동정되어 있었다. 또한 마의 어린 개체가 둥근마로 오동정되어 있었으며, 일본에서 채집된 표본만이 둥근마로 확인되었을 뿐이다. 따라서 본 연구에서는 국화마와 둥근마의 자생지를 확인할 수 없었고, 따라서 두 종의 국내 분포 여부에 대해서는 재고되어야 할 것으로 판단된다.
Fig 1.
The types of root of Korean Dioscorea L. A. Vertical tuberous root; B. Horizontal rhizome 
Fig 2.
The types of bulbil of Korean Dioscorea L. A. Bulbil; B. Non-Bulbil 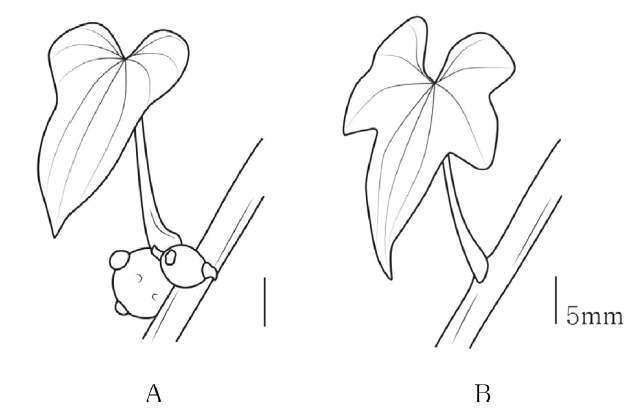
Fig 3.
The petiole base of Korean Dioscorea L. A. Protrude papillae; B. Non-protrude papillae. 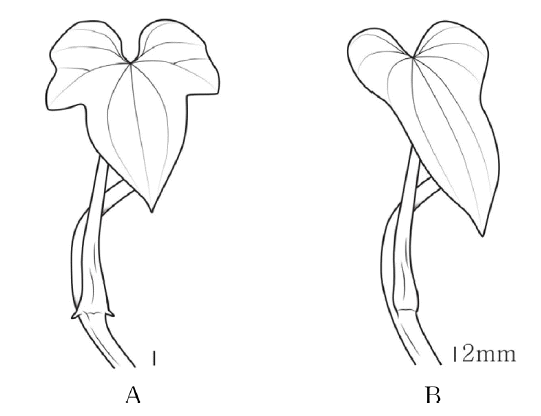
Fig 4.
The shapes of leaf of Korean Dioscorea L. A. D. polystachya; B. D. japonica; C. D. nipponica; D. D. quinqueloba; E. D. tenuipes; F. D. tokoro. 
Fig 5.
The comparison of leaf of D. nipponica and D. quinqueloba. A. D. nipponica; B. D. quinqueloba (1: length of middle-lobe; 2: width of middle-lobe; 3: length of First-lobe; 4: width of First-lobe; 5: length of Second-lobe; 6: width of Second-lobe; 7: First-angle; 8: Second-angle). 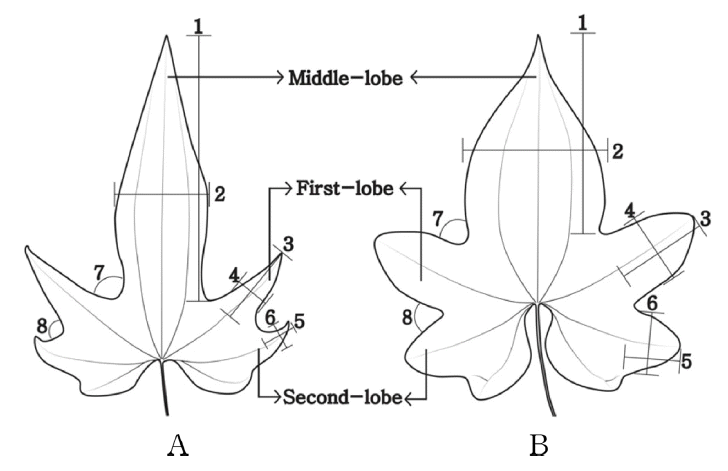
Fig 6.
The inforescence of Korean Dioscorea L. A. D. polystachya; B. D. japonica; C. D. nipponica, D. quinqueloba; D. tenuipes; D. tokoro. 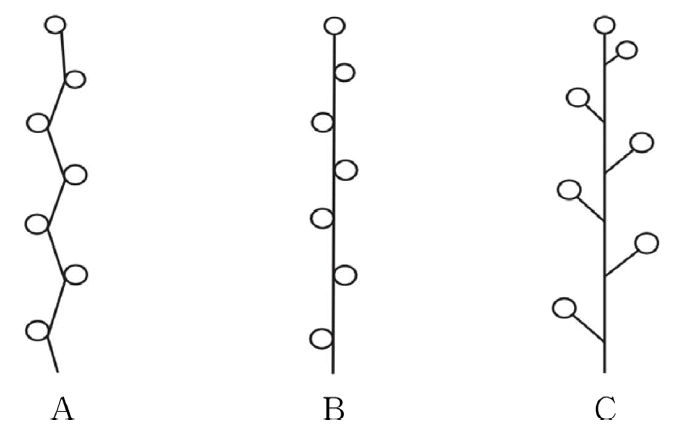
Fig 7.
The shapes of tepal of Korean Dioscorea L. A. D. polystachya, D. japonica; B. D. nipponica, D. quinqueloba; C. D. tokoro; D. D. tenuipes. 
Fig 8.
The shape of flower of Korean Dioscorea L. Aa. Ab. D. polystachya; Ba. Bb. D. japonica; Ca. Cb. D. nipponica; Da. Db. D. quinqueloba; Ea. Eb. D. tenuipes; Fa. Fb. D. tokoro. 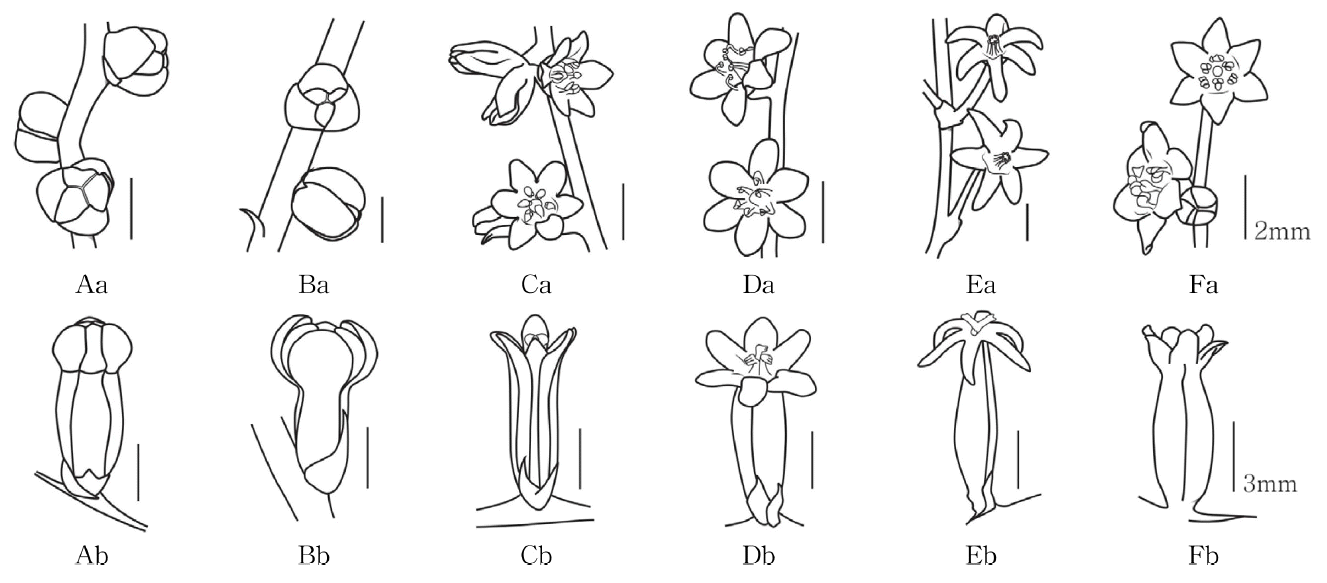
Fig 9.
The shapes of stamen and pistil of Korean Dioscorea L. A. Stamen (a, b. D. polystachya; c, d. D. japonica; e, f. D. nipponica; g. D. quinqueloba; h. D. tenuipes; i. D. tokoro), B. Pistil (a. D. polystachya; b. D. japonica; c. D. nipponica; d. D. quinqueloba; e. D. tenuipes; f. D. tokoro). 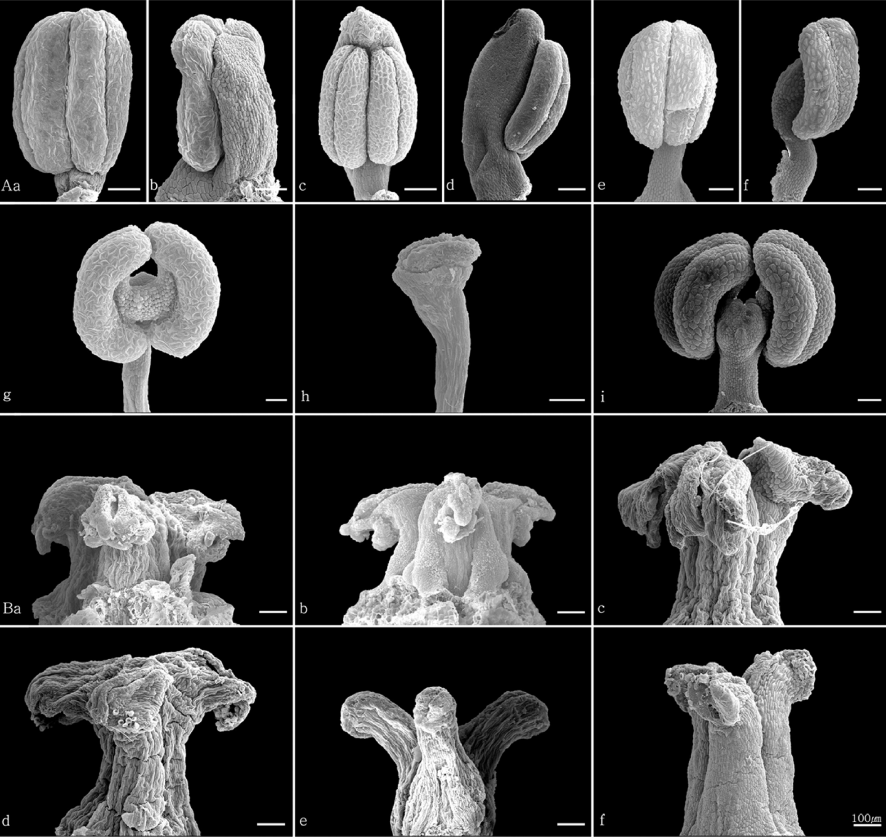
Fig 10.
The shapes of fruit and seed of Korean Dioscorea L. Aa. Ab. D. polystachya; Ba. Bb. D. japonica; Ca. Cb. D. nipponica; Da. Db. D. quinqueloba; Ea. Eb. D. tenuipes; Fa. Fb. D. tokoro. 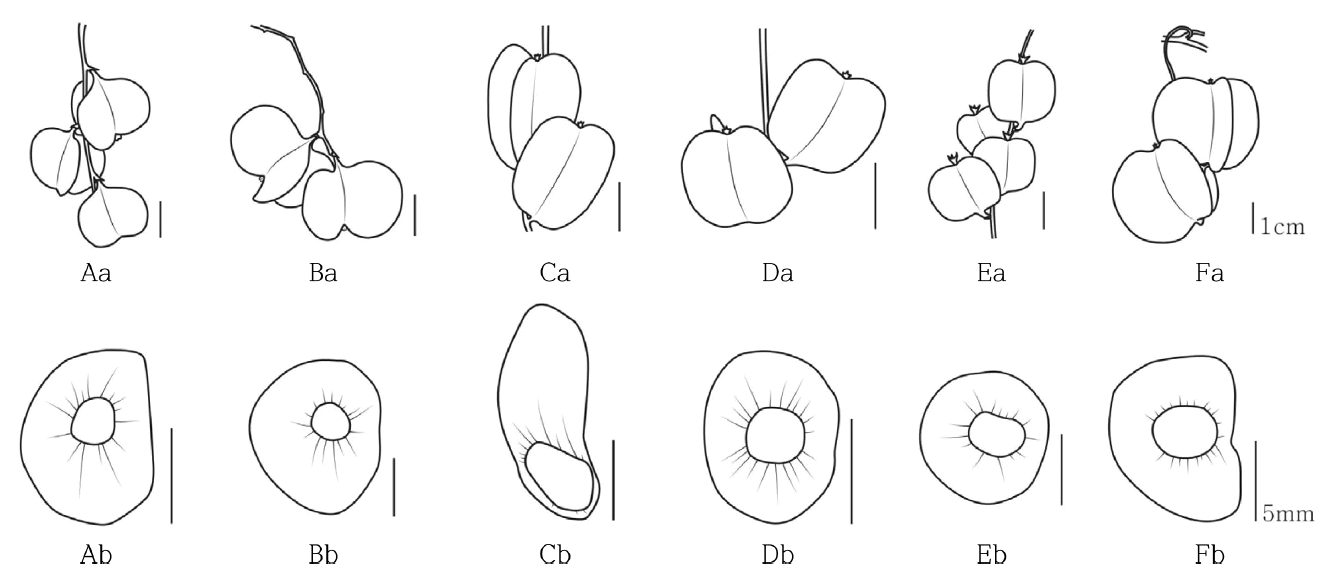
Fig 11.
Distribution map of Korean Dioscorea L. D. polystachya; D. japonica; D. nipponica; D. quinqueloba; D. tenuipes; D. tokoro. 
Table 1.
Measurement on external morphology of genus Dioscorea in Korea.
|
Underground part
|
Inflorescence
|
Bulbil
|
Petiole base
|
Leaf
|
Periacth
|
Fruit
|
Seed
|
|
Type |
Male |
Female |
Present or Absent |
Form |
Shape |
Type |
Tepal color |
Tepal shape |
Shape |
Shape |
Wing position |
|
D. polystachya
|
tuberous |
spike |
spike |
Present |
non-protuberance |
deltate |
closed |
white |
widely-ovate |
obovate-globose |
domed |
all round |
|
D. japonica
|
tuberous |
spike |
spike |
Present |
non-protuberance |
narrowly-triangular |
closed |
white |
widely-ovate |
broadly ovate |
globose |
all round |
|
D. nipponica
|
rhizomatous |
raceme |
raceme |
Absent |
non-protuberance |
deltate-ovate |
campaniform |
yellowish |
obovate |
Oblong |
oblong |
one side |
|
D. quinqueloba
|
rhizomatous |
raceme |
raceme |
Absent |
protuberance |
deltate-ovate |
campaniform |
orange |
obovate |
broadly ovate |
globose |
all round |
|
D. tenuipes
|
rhizomatous |
raceme |
raceme |
Absent |
protuberance |
triangular-cordiform |
reflexed |
yellowish-green |
linear |
broadly ovate |
globose |
all round |
|
D. dokoro
|
rhizomatous |
raceme |
raceme |
Absent |
non-protuberance |
cordiform |
campaniform |
ivory |
ovate |
broadly ovate |
domed |
all round |
|
Leaf (cm)
|
|
Male flower (mm)
|
|
Petiole
|
Blade
|
*length |
*width |
Outer tepal
|
Inner tepal
|
|
*length |
*length |
*width |
L/W |
*length |
*width |
*length |
*width |
|
D. polystachya
|
2.6 (3.4) 4.0 |
4.3 (5.6) 6.4 |
4.2 (4.8) 5.6 |
1.1 |
1.8 (1.9) 2.3 |
1.7 (1.9) 2.3 |
1.5 (1.7) 2.0 |
1.1 (1.4) 1.8 |
1.1 (1.3) 1.6 |
0.9 (1.1) 1.3 |
|
D. japonica
|
1.7 (3.4) 5.7 |
6.4 (8.7) 13.7 |
2.9 (4.3) 6.7 |
2.0 |
2.1 (2.4) 2.7 |
1.5 (1.8) 2.2 |
2.2 (2.4) 2.6 |
1.4 (1.7) 2.0 |
1.4 (1.6) 1.8 |
1.1 (1.4) 1.6 |
|
D. nipponica
|
3.3 (5.8) 8.9 |
6.1 (8.3) 11.4 |
4.6 (6.9) 10.4 |
1.1 |
2.2 (2.6) 2.9 |
2.2 (2.5) 2.9 |
2.0 (2.2) 2.5 |
1.3 (1.5) 1.6 |
1.9 (2.2) 2.5 |
1.1 (1.4) 1.6 |
|
D. quinqueloba
|
3.8 (4.9) 7.0 |
8.1 (9.6) 12.6 |
7.1 (8.7) 10.9 |
1.1 |
0.7 (1.0) 1.2 |
3.1 (3.6) 4.3 |
1.7 (1.8) 2.0 |
0.8 (0.9) 1.1 |
1.5 (1.6) 1.9 |
0.7 (0.8) 1.0 |
|
D. tenuipes
|
2.0 (3.1) 4.8 |
4.2 (7.2) 8.9 |
2.8 (4.4) 5.7 |
1.6 |
1.1 (1.3) 1.4 |
3.4 (3.8) 4.2 |
1.3 (1.6) 1.9 |
0.4 (0.5) 0.6 |
1.2 (1.5) 1.9 |
0.4 (0.5) 0.6 |
|
D. dokoro
|
5.3 (6.7) 9.3 |
5.9 (7.2) 8.5 |
4.3 (5.7) 7.7 |
1.2 |
1.6 (1.9) 2.1 |
3.3 (3.6) 4.2 |
2.2 (2.3) 2.5 |
1.2 (1.3) 1.5 |
2.1 (2.2) 2.5 |
1.1 (1.3) 1.5 |
|
Male flower (mm)
|
|
Female flower (mm)
|
|
Stamen
|
Anther
|
*length |
*width |
Outer tepal
|
Inner tepal
|
|
*length |
*width |
*length |
*width |
*length |
*width |
*length |
*width |
|
D. polystachya
|
0.9 (1.1) 1.2 |
0.4 (0.4) 0.5 |
0.4 (0.5) 0.6 |
0.2 (0.25) 0.3 |
1.2 (1.5) 1.9 |
2.0 (2.3) 2.5 |
1.4 (1.5) 1.6 |
1.3 (1.4) 1.6 |
1.0 (1.4) 1.6 |
1.1 (1.3) 1.5 |
|
D. japonica
|
0.7 (0.8) 0.9 |
0.2 (0.3) 0.3 |
0.5 (0.5) 0.6 |
0.2 (0.2) 0.2 |
1.2 (1.7) 1.9 |
1.6 (2.1) 2.3 |
1.4 (1.5) 1.7 |
1.3 (1.4) 1.5 |
1.1 (1.2) 1.3 |
1.3 (1.5) 1.6 |
|
D. nipponica
|
0.6 (0.7) 0.8 |
0.1 (0.2) 0.3 |
0.4 (0.5) 0.6 |
0.1 (0.2) 0.3 |
1.2 (1.4) 1.7 |
2.3 (2.8) 3.3 |
1.4 (1.6) 1.8 |
0.6 (0.8) 1.0 |
1.4 (1.5) 1.7 |
0.6 (0.8) 1.0 |
|
D. quinqueloba
|
0.6 (0.7) 0.8 |
0.1 (0.1) 0.2 |
0.2 (0.3) 0.4 |
0.1 (0.1) 0.2 |
1.5 (1.9) 2.2 |
3.3 (3.9) 4.6 |
1.7 (2.0) 2.5 |
1.1 (1.3) 1.6 |
1.5 (1.8) 2.4 |
1.0 (1.2) 1.6 |
|
D. tenuipes
|
0.6 (0.6) 0.7 |
0.7 (0.8) 0.1 |
0.2 (0.3) 0.3 |
0.1 (0.1) 0.1 |
0.7 (0.9) 1.1 |
3.3 (3.6) 4.2 |
1.6 (1.8) 2.0 |
0.3 (0.4) 0.5 |
1.6 (1.8) 2.0 |
0.3 (0.4) 0.5 |
|
D. dokoro
|
0.7 (0.8) 0.9 |
0.1 (0.1) 0.2 |
0.5 (0.5) 0.6 |
0.2 (0.2) 0.3 |
2.0 (2.3) 2.6 |
2.4 (2.8) 3.2 |
1.5 (1.7) 2.0 |
0.9 (1.0) 1.2 |
1.4 (1.7) 2.0 |
0.8 (1.0) 1.2 |
|
Female flower (mm)
|
Fruit (mm)
|
Seed (mm)
|
Seed Wing (mm)
|
|
Pistil
|
*length |
*width |
*length |
*width |
*length |
*width |
|
*length |
*width |
|
D. polystachya
|
0.3 (0.4) 0.5 |
0.8 (0.9) 1.0 |
9.7 (14.4) 15.5 |
16.6 (17.1) 17.9 |
3.5 (3.9) 4.3 |
3.0 (3.5) 4.1 |
7.9 (9) 10.5 |
11.9 (13.6) 16.5 |
|
D. japonica
|
0.4 (0.4) 0.5 |
0.8 (0.9) 1.0 |
15.1 (17.1) 18.5 |
24.6 (27.4) 29.5 |
3.6 (4.2) 4.8 |
3.7 (4.2) 4.6 |
11.6 (13.5) 14.9 |
14.4 (15.8) 17.3 |
|
D. nipponica
|
1.2 (1.4) 1.6 |
0.9 (1.0) 1.1 |
18.1 (19.9) 22.2 |
15.9 (17.8) 19.0 |
4.3 (5.6) 6.6 |
2.5 (3.4) 4.2 |
11.3 (13.6) 15.8 |
5.3 (6.6) 7.5 |
|
D. quinqueloba
|
0.6 (0.7) 0.7 |
0.8 (0.9) 1.1 |
12.3 (13.1) 13.9 |
14.6 (15.9) 16.8 |
3.7 (4.5) 4.9 |
3.0 (3.5) 4.1 |
7.8 (8.2) 8.7 |
9.4 (10.4) 11.3 |
|
D. tenuipes
|
0.4 (0.5) 0.6 |
0.9 (0.1) 1.1 |
15.1 (16.0) 17.2 |
19.2 (20.0) 20.9 |
4.5 (5.0) 5.7 |
3.4 (4.1) 4.6 |
7.5 (9.4) 11.2 |
9.1 (10.3) 11.4 |
|
D. dokoro
|
0.7 (0.8) 0.9 |
0.6 (0.8) 0.9 |
21.5 (23.5) 25.1 |
25.5 (27.6) 30.5 |
4.5 (5.2) 5.8 |
3.8 (4.2) 4.5 |
8.8 (9.8) 11.2 |
14.1 (15.1) 16.1 |
Table 2.
The comparison between of D. nipponica and D. quinqueloba.
|
Leaf
|
|
Middle-lobe
|
First-lobe
|
|
apex |
*length |
*width |
apex |
*length |
*width |
|
D. nipoonica
|
acute |
4.1 (5.4) 6.8 |
2.5 (3.3) 4.0 |
acute |
1.0 (2.1) 4.1 |
1.0 (1.4) 1.9 |
|
D. quinqueloba
|
acute |
2.7 (5.3) 7.5 |
2.5 (3.9) 5.8 |
obtuse |
1.0 (2.0) 3.1 |
1.3 (2.0) 2.6 |
|
Leaf
|
|
Second-lobe
|
Angle (°)
|
|
apex |
*length |
*width |
first-angle |
second-angle |
|
D. nipoonica
|
acute |
0.5 (1.6) 2.8 |
1.0 (1.4) 1.8 |
50 (57) 65 |
52 (77) 110 |
|
D. quinqueloba
|
obtuse |
0.8 (1.3) 1.7 |
1.2 (1.7) 2.5 |
40 (63) 83 |
50 (70) 110 |
Literature Cited
Beccari, O. 1870. Nuovo Giornale Botanico Italiano Publicato da O. Beccari. Stabilimento di G. Pellas, Firenze. V. 2. 12−13.
Burkill, I. H. 1960. The organography and the evolution of the Dioscoreaceae, the family of the yams. Botanical Journal of the Linnean Society 56: 319-412.  Coursey, D. G. 1969. The future of the yam in world agriculture: an appraisal of some problems. Tropical Root and Tuber Crops Newsletter 2: 4-8.
Decaisne, J. 1854. Revue Horticole. Librairie agricole de la maison rustique, Paris. Ser. IV, iii. P. 243.
Ding, Z. and Gilbert, M. G.. 2000. Dioscoreaceae. Wu, Z.. Raven, P. H. (eds.), Flora of China. Flagellariaceae through Marantaceae. Science Press; Beijing. Missouri Botanical Garden Press; St. Louis. 24: 276-296.
Engler, A. and Prantl, K.. 1897. Die natürlichen Pflanzenfamilien V. 2(5). Wilhelm Engelmann, Leipzig, Leipzig. 80-87.
Gao X, X.. Zhu, Y. P.. Wu, B. C.. Zhao, Y. M.. Chen, J. Q. and Hang, Y. Y.. 2008. Phylogeny of Dioscorea sect. Stenophora based on the chloroplast matK, rbcL and trnL-F sequences. Journal of Systematics and Evolution 46: 315-321.
Gawande, P. A.. Nimbhorkar, N. V. and Thakare, V.. 2015. Distribution and morphodiversity analysis of genus Dioscorea from India with special reference to Satpura Hilly Ranges and Western Ghat of Maharashtra state, India. International Journal of Basic and Applied Sciences 4: 146-150.
Hsu, K. M. Tsai, J. L.. Chen, M. Y.. Ku, H. M. and Liu, S. C.. 2013. Molecular phylogeny of Dioscorea (Dioscoreaceae) in East and Southeast Asia. Blumea 58: 21-27. Im, R. J. 2000. Flora Koreana. Vol. 9. The Science and Technology Publishing House, Pyongyang, Korea. 217-221.
Kang, F. H. Lee, C. S. and Oh, Y. C.. 1992. A systematic study of sect. Enantiophyllum and Stenophora taxa (Dioscorea, Dioscoreaceae) in Korea. Korean Journal of Plant Taxonomy 22(4): 255-278.
Knuth, C. S. 1850. Dioscorineae. In: Enumeratio Plantarum. V. 5. Sumtibus J. G. Cottae, Stutgardiae et Tubingae. 323-456.
Lee, T. B. 2003. Coloured Flora of Korea. Vol.. Hayangmunsa, Seoul. 727-731 (in Korean).
Lee, Y. N. 2006. New Flora of Korea. Vol.. Kyohak Publising Co, Seoul. 473-475 (in Korean).
Li, H. L. Liu, T. S.. Huang, T. C.. Koyama, T. and De, C. E.. 1978. Flora of Taiwan. Epoch Publishing Co., Ltd, Vol. 5: 99-109.
Linnaeus, C.. 1753. Diosocrea In: Species Plantarum. V. 2.. Impensis Laurentii Salvii, Holmiae. 1032-1034.
Maekawa, F.. Hara, H. and Tuyama, T.. 1962. Makino’s New Illustrated Flora of Japan. Hohuryukan. Co. Ltd., Tokyo. 881-884.
Matsuno, G.. 1918. Dioscorea in Japan. Journal of Japanese Botany 1: 282-286.
Mori, T.. 1921. An Enumeration of Plants hitherto Known from Corea. The government of Chosen, Seoul. 96-97.
Nakai, T.. 1911. Flora Koreana. Pars secunda. The Journal of the College of Science. Imperial University of Tokyo, 31: 235 pp.
Nakai, T.. 1952. A synoptical Sketch of Korean Flora. Bulletin of the National Science Museum; Tokyo. 31: 148.
Oh, Y. C. Lee, C. S. and Lee, H. J.. 1995. A systematic study on Dioscorea sect. Enantiophyllum and Stenophora in Korea: Microstructure and chemistry. Korean Journal of Plant Taxonomy 25(1): 25-49.
Oh, Y. C. Chang, C. S. and Lee, K. A.. 1996. Morphometric analyses of Dioscorea. Korean Journal of Plant Taxonomy 26(2): 125-140.
Oh, Y. C. and Lee, C. S.. 2007. The Genera of Vascular Plants of Korea. Academy Publishing Co., Seoul. 1336-1338 (in Korean).
Ohwi, J.. 1984. Flora of Japan. Smithsonian Institution, Washington DC. 313-315.
Palibin, J.. 1901. Conspectus Florae Koreae. Pars tertia. Trudy Imperatorskago S.-Peterburgskago Botaniceskago Sada. 19: 103-151.
Park, M. K.. 1949. An Enumeration of Korean Plants. 330-331 (in Korean).
Tominari, T.. Wanteri, S.. Kitamura, S.. Ohwi, J. and Satake, Y.. 1984. Wild Flowers of Japan. Heibonsya, Tokyo. 1-56.
Turczaninow, P. K. N. S. 1837. Bulletin de la Societe Imperiale des Naturalistes de Moscou. 7: 158 pp.
Wilkin, P.. Schols, P.. Chase, M. W.. Chayamarit, K.. Furness, C. A.. Huysmans, S.. Rakotonasolo, F.. Smets, E. and Thapyai, C.. 2005. A Plastid gene phylogeny of the yam genus, Dioscorea: Roots, Fruits and Madagascar. Systematic Botany 30: 736-749.
|
|











