/home/virtual/kjpt/journal//../xmls/kjpt-51-1-1.xml
LEE, KIM, KIM, KWON, YANG, CHO, KIM, LEE, MAKI, and KIM: Symplocarpus koreanus (Araceae; Orontioideae), a new species based on morphological and molecular data
Abstract
Symplocarpus koreanus (Araceae; Orontioideae) from Korea is described as new. Symplocarpus koreanus has long been considered to be conspecific with S. renifolius in Japan, but phylogenetic, morphological, and cytological data indicate its taxonomic distinction. Compared to S. renifolius, S. koreanus has a much smaller spathe and more spherical spadix with fewer, smaller flowers. Previous phylogenetic studies also suggested that S. koreanus is more closely related to nonthermogenic S. nipponicus than to S. renifolius in Japan. Like its nonthermogenic sister species, S. nipponicus, in Korea and Japan, S. koreanus is diploid (2n = 2x = 30), while S. renifolius in Japan is tetraploid (2n = 4x = 60). A detailed species description, geographical distribution, major morphological differences between the species and a dichotomous key to the species in eastern Asia are provided.
Keywords: Araceae, Orontioideae, Symplocarpus koreanus, Korean endemic, Korean skunk cabbage
Symplocarpus Salisb. ex W. P. C. Barton (Araceae, Orontioideae), a genus with disjunct distribution, represents one of a number of eastern Asian-eastern North American examples ( Mayo et al., 1997). Symplocarpus includes five species, all but one occurring in eastern Asia. Symplocarpus foetidus (L.) Salisb. ex W. P. C. Barton is the only species in eastern North America, occurring from Nova Scotia, southern Quebec, and Minnesota to North Carolina and Tennessee. In eastern Asia, two species, S. renifolius Schott ex Tzvelev and S. nipponicus Makino, occur broadly throughout Japan, Korea, northeast China and the Russian Far East ( Ohwi, 1984; Tsvelev, 1996; Ko, 2007; Li et al., 2010; Murata, 2016), while two recently described species, S. nabekuraensis Otsuka & K. Inoue and S. egorovii N. S. Pavlova & V. A. Nechaev, occur narrowly in northern Nagano Prefecture in central Japan and the Primorye region in Russia, respectively ( Otsuka et al., 2002; Pavlova and Nechaev, 2005). Two intercontinental disjunct sister species, S. foetidus in eastern North America and S. renifolius in eastern Asia, share several characteristics, such as flowering in early spring before the emergence of leaves, fruit ripening in the fall of the same year and exothermic spadices ( Wada and Uemura, 2000; Ito et al., 2004). Symplocarpus egorovii is similar to S. renifolius and S. foetidus, but differs in its yellowish white spathe and distinct habitat in coniferous forests ( Pavlova and Nechaev, 2005). Symplocarpus nabekuraensis can be distinguished from its congeneric species in Japan based on leaf shape and peduncle length ( Otsuka et al., 2002). While S. nipponicus and S. egorovii are diploid (2 n = 2 x = 30), S. renifolius, S. foetidus, and S. nabekuraensis are tetraploid (2 n = 4 x = 60) ( Sokolovskaya and Probatova, 1985; Iwatsubo and Otsuka, 2005; Marhold, 2008; Probatova et al., 2012). The three tetraploid species are known to have an exothermic spadix, whereas the diploid S. nipponicus has a non-exothermic spadix ( Knutson, 1974; Wada and Uemura, 1994; Ito et al., 2004; Otsuka et al., 2011).
In a phylogeny of the subfamily Orontioideae based on extensive sampling in eastern Asia and eastern North America, we recognized several novel features ( Nie et al., 2006; Lee et al., 2019). Two major lineages of Symplocarpus were revealed: one lineage included S. foetidus, S. nabekuraensis and S. renifolius (Japan); the other included S. nipponicus, S. egorovii and S. renifolius (Korea). We further confirmed that S. renifolius in Korea was diploid (2 n = 30), while S. renifolius in Japan and S. foetidus were tetraploid ( Lee et al., 2019). The large scale phylogeny of Symplocarpus and chromosome numbers suggested that S. renifolius in Korea represented a distinct taxon. In this study, we compared several morphological characteristics between populations of S. renifolius in Japan and Korea. Based on the molecular, morphological, and cytological evidence, we here describe the populations of S. renifolius in Korea as a distinct taxon, S. koreanus.
Taxonomic Treatment
Symplocarpus koreanus J. S. Lee, S. H. Kim & S. C. Kim, sp. nov. ( Fig. 1)—TYPE: Korea. Gangwon-do: Chuncheon-si, Sabuk-myeon, Goseong-ri, Sayeogol-gil, Yonghwasan Mountain, N 38 00.918, E 127 45.286, elev. 333 m, 21 Mar 2020, S. C. Kim 200321500 (holotype: SKK; isotype: KB).
Herbs, perennial, seasonally dormant. Roots contractile, string-like, 1.6–5.3 mm in diam. Rhizome hypogenous, erect, 10–27 mm in diameter. Leaves in a rosette, appearing after flowers, 2–5; petiole basally sheathing, sulcate, purplish striped, 5.8–17 cm × 1.9–10.2 mm; leaf blade subcordate or cordate-ovate, 7–24 × 4.6–19 cm, yellowish green or green, base cordate or reniform, slightly uneven, margin entire, apex acute, acuminate, or obtuse; venation reticulate, midvein prominent, primary lateral veins 5–7 on each side of midvein, arching and branching apically, merging with inconspicuous marginal veins, lowermost veins running into basal lobes. Inflorescences 1–3, in a sympodium, appearing before leaves, 29–62 flowered; peduncle erect, partly below ground, shortly exserted above ground, terete, 3.5–21 cm × 0.3–2.1 mm, yellowish green or purplish tinged. Spathe hood-like, convolute at base, fleshy, 6–11 × 2.8–5.8 cm, yellowish brown or yellowish green, usually dark purple spotted or striped, apex 2-keeled, acute or acuminate, twisted or incurved, not persisting in fruit. Spadix subglobose or ellipsoid, 9.3–17 × 7.4–15 mm, stipitate; stipe cylindrical, 2–9.3 × 2–4.4 mm. Flowers bisexual, 2.7–5.5 × 2.3–5 mm; tepals 4, imbricate, yellow; stamens 4, free; filaments flattened, connective slender; anthers 2-locular, basifixed, yellow, dehiscing longitudinally; ovary 1-locular; ovule 1; style cylindrical, truncate or capitate. Infructescence globose or broadly ellipsoid, spadix 2.7–3.3 × 2.4–3 cm, stipe 10.8–19 × 4.5–6.3 mm. Fruit a berry, embedded in enlarged spongy spadix, initially greenish or dark purple, becoming blackish brown at maturity, ripening below ground in summer of same year. Seed brown, somewhat irregular in shape, 4.6–11.5 × 4.8–13.7 mm, smooth. Chromosome number: 2n = 30.
Etymology: The specific epithet, koreanus, is based on the name of the country of origin, Korea.
Vernacular name: Han-guk-an-jeun-bu-chae (한국앉은부채).
English colloquial name: Korean skunk cabbage.
Phenology: Flowering late February to April.
Chromosome number: 2 n = 2 x = 30. 2C value range = 2.72–6.25 pg. ( Fig. 2)
Distribution and ecology: Symplocarpus koreanus is restricted to the Korean Peninsula (North Korea, including Hamgyeongnam-do and South Korea, including Gyeonggi-do, Gangwon-do, Chungcheongbuk-do, Jeollabuk-do, Gyeongsangbuk -do, and Gyeongsangnam-do). It typically occurs in shaded, wet places, along streams and on moist rocky hillsides in mixed deciduous forests from ca. 100 to 750 m elevation.
Additional specimens examined: KOREA. Gyeonggi-do: Gapyeong-gun, Buk-myeon, Baekdun-ri, Yeoninsan Mt., 3 May 2013, W. B. Lee, NIBRVP0000517078 (KB); Gapyeong-gun, Oeseo-myeon, Hwayasan Mt., 26 Jun 2007, W. K. Paik, VP-KB-377062-0173 (KB); Gapyeong-gun, Sang-myeon, Haenghyeon-ri, Chungnyeongsan Mt., 29 Mar 2016, G. H. Nam et al., NIBRVP0000550794 (KB); 31 Mar 2012, J. H. Kim et al., NIBRVP0000355001 (KB); Gapyeong-gun, Seorak-myeon, Yumyeongsan Mt., 4 Apr 2008, B. G. Kwon, NIBRVP0000532404 (KB), G. Y. Chung, NIBRVP0000197125 (KB); Gwacheon-si, Makgye-dong, Cheonggyesan Mt., 5 Apr 2012, Y. S. Kim, 021215 (SKK); Gwangju-si, Taehwasan Mt., 19 Apr 2013, K. I. Heo et al., 021210 (SKK); Hanam-si, Baealmi-dong, Geomdansan Mt., 3 Aug 2007, J. O. Hyun et al., NIBRVP0000111433 (KB); 11 Sep 2015, M. S. Cho et al., 035869 (SKK); Namyangju-si, Hwado-eup, Cheonmasan Mt., 15 Apr 2007, W. K. Paik, VP-KB-377061-0133 (KB); 22 Mar 2013, Song et al., NIBRVP0000464822 (KB); Namyangju-si, Jinjeop-eup, Gwangreung, 24 May 1954, T. G. Joo, 021191 (SKK); Jun 1965, T. H. Chung, 021193 (SKK); Namyangju-si, Onam-eup, Onam-ri, Cheonmasan Mt., 6 Apr 2009, G. H. Nam et al., NIBRVP0000206699, NIBRVP0000206700 (KB); Namyangju-si, Sudong-myeon, Chukryeongsan Mt., 28 Mar 1999, S. P. Hong and K. W. Park, NIBRVP0000102296 (KB); Paldalsan Mt., 31 May 1967, T. B. Lee, 00072835 (SNUA); Sudong-myeon, Namyangju-si, Chukryeongsan Mt., 5 Apr 2005, J. Kim et al., 021204, 021224 (SKK); M. H. Lim et al., 021232, 21203 (SKK); N. I. Gong et al., 21202 (SKK); S. R. Lee et al., 021199 (SKK), T. E. Park et al., 021200 (SKK); 6 Feb 2007, S. T. Lee et al., 021201 (SKK); 8 May 1993, K. R. Lim et al., 021192 (SKK); 9 Apr 2010, S. C. Kim et al., 021229 (SKK); 27 Apr 2007, K. I. Heo et al., 021225, 021226, 021227, 021228, 021234 (SKK); 27 Mar 1992, S. T. Lee et al., 021195, 021196 (SKK); 30 Mar 2002, S. T. Lee et al., 021233 (SKK). Gangwon-do: Cheongnyeongpo, 13 Aug 2004, J. M. Chung et al., 04080073 (KH); Cheorwon-gun, Geunnam-myeon, Gwangdeoksan Mt., 12 May 1997, S. P. Hong and H. S. Choi, NIBRVP0000102297 (KB); Chuncheon-si, Sabuk-myeon, Jipdarigol Recreation Forest, 30 May 2013, S. C. Kim et al., 021208, 021209 (SKK); Chuncheon-si, Sabuk-myeon, Yonghwasan Mt., 5 Apr 2012, K. I. Heo et al., 021211 (SKK); 30 May 1998, D. M. Choi et al., 0003644 (HUH); 21 Mar 2020, S. C. Kim et al., 200321001, 200321002, 200321003, 200321004, 200321005, 200321006, 200321007, 200321008, 200321009, 200321010, 200321011, 200321012, 200321013, 200321014, 200321015, 200321016, 200321017, 200321018, 200321019, 200321020, 200321021, 200321022, 200321501, 200321502, 200321503, 200321504 (SKK); 13 Jun 2020, S. C. Kim et al., 200613001, 200613002, 200613003, 200613004, 200613005, 200613006, 200613007, 200613008, 200613009, 200613010, 200613011, 200613012, 200613013 (SKK); Chuncheon-si, Seo-myeon, Deokduwon-ri, Samaksan Mt., 1 Jul 2012, K. I. Heo et al., 021216, 021217,021231 (SKK); 30 Mar 2013, S. C. Kim et al., 021206 (SKK); Chuncheon-si, Sindongmyeon, Jeongjok-ri, Geumbyeongsan Mt., 19 Jun 2007, G. E. Yu, 063114 (KWNU); 1 Jul 2012, K. I. Heo et al., 021221 (SKK); 30 Mar 2013, S. C. Kim et al., 021207 (SKK); Deungseon Falls, 27 May 1995, H. G. Lee et al., 0003645 (HUH); Donghaesi, Bugok-dong, Mita Temple, 26 Apr 2011, G. H. Nam and W. J. Jeong, NIBRVP0000284290 (KB); Gangneung-si, Wangsan-myeon, Gangneung-si, Hwaranbong Peak, 30 Apr 2009, J. H. Kim and H. J. Kim, NIBRVP0000318582 (KB); Garisan Mt., 15 Jun 2011, K. O. Yoo et al., koyoo0118 (KH); Hugog Mineral Spring, 4 Jun 1999, C. S. Chang, 00070596 (SNUA); Hwacheon-gun, Baekjeoksan Mt., 24 May 2000, K. C. Yang and J. D. Jung, NIBRVP0000102304, NIBRVP0000102305 (KB); Hwacheon-gun, Sanae-myeon, Gwangdeok-ri, Gwangdeoksan Mt., 7 Apr 2007, S. T. Lee et al., 021198, 021222 (SKK); Apr 2009, G. H. Nam et al., NIBRVP0000206708 (KB); 7 Aug 2013, S. T. Lee et al., 021205 (SKK); 7 May 2010, K. I. Heo, 021230 (SKK); 15 Apr 2006, J. H. Lee et al., 021223 (SKK); 18 May 2012, K. I. Heo et al., 021212, 021213, 021214, 021218, 021219, 021220 (SKK); 7 Apr 2009, G. H. Nam et al., NIBRVP0000206709 (KB); Hwacheon-gun, Sangseo-myeon, Baekjeoksan Mt., 3 Aug 2000, J. H. Kim and D. K. Kim, NIBRVP0000102307 (KB) Chungcheongbuk-do: Danyang-gun, Gagok-myeon, Sobaecksan Mt., 17 May 1999, C. W. Park et al., EN99SNU315 (KB); 20 Apr 2007, G. Y. Chung, NIBRVP0000121631 (KB); Dutasan Mt., 25 May 2002, B. U. Oh et al., 04206 (CBU); Gaseopsan Mt., 30 Apr 2000, S. H. Paeng et al., 04204 (CBU); Geumdangsan Mt., 22 May 1999, C. S. Chung et al., 04202 (CBU); Guryongsan Mt., 17 May 1997, B. U. Oh et al., 07317 (CBU); Ingyeongsan Mt., 21 May 2011, B. U. Oh et al., 22620 (CBU); Seondosan Mt., 31 May 1997, B. U. Oh et al., 08908 (CBU); Sosokisan Mt., 13 May 2005, B. H. Oh et al., Eumseonggun (Sosogisan)-0505 (KH). Jellabuk-do: Jangsu-gun, Geunnam-myeon, Jangan-ri, 21 Sep 1997, B. Y. Sun and C. H. Kim, EN98JNU 464 (KB); Jangsu-gun, Gyenam-myeon, Jangansan Mt., 19 Jun 2009, J. K. Ahn et al., NIBRVP0000266477, NIBRVP0000266707 (KB); 19 May 2007, B. Y. Sun, NIBRVP0000128343 (KB); Jinan-gun, Jucheon-myeon, Daebul-ri, Unjangsan Mt., C. H. Kim and S. H. Lee, NIBRVP0000537859 (KB); Sunchang-gun, Gurim-myeon, Ansim-gil, Hoemunsan National Recreation Forest, 14 Mar 2014, S. H. Kim et al., 2014314500 (SKK). Gyeongsangbuk-do: Bibongsan Mt., 6 Aug 2000, S. W. Lee et al., 218 (KNU); Gallasan Mt., 6 Jun 2008, M. S. Park, ANH-000014010 (ANH); Jachosan Mt., 13 May 2010, G. Y. Chung et al., ANH-100513125 (ANH); Joosan Mt., 20 May 1999, D. R. Choi and S. G. Kwon, 00072844 (SNUA); Maengdongsan Mt., 28 Aug 2006, G. Y. Jeong, Jeong210 (KH); Muposan Mt., 2 Oct 2010, G. Y. Chung et al., ANH-000012103 (ANH); Podosan Mt., 7 Jul 2007, G. Y. Chung et al., ANH-0001504 (ANH). Gyeongsangnam-do: Geochang-gun, Buksang-myeon, Deokyusan Mt., 31 May 2006, B. Y. Sun, NIBRVP0000119643 (KB).
Note: Symplocarpus koreanus has long been considered to be conspecific with S. renifolius in Japan ( Appendix 1). A recent phylogenetic study based on extensive sampling revealed that S. renifolius in Korea, now described as S. koreanus, is more closely related to S. nipponicus in Korea and Japan than it is to S. renifolius in Japan and the Russian Far East ( Lee et al., 2019).
Morphologically, S. koreanus is most similar to S. renifolius in Japan, including in life history traits, perhaps sharing their most recent common ancestor. However, S. koreanus has a much smaller and more spherical spadix with fewer, smaller flowers than S. renifolius ( Tables 1, 2). The average spadix length of S. koreanus is 12 mm, while that of S. renifolius is 22 mm. In addition, the average spadix width of S. koreanus is 10 mm, whereas that of S. renifolius is 15 mm. The ratio between spadix length (L) and width (W) is also different: 1.2 and 1.4 for S. koreanus and S. renifolius, respectively. Lastly, S. koreanus has many fewer flowers per spadix than S. renifolius: average of 40 versus 79. Thus, the spadix of S. koreanus in its size, shape and flower number is more similar to the spadix of S. nipponicus than to S. renifolius. We utilized flow cytometry to estimate ploidy level between S. koreanus and S. renifolius (see comprehensive review and references in Kron et al., 2007). Briefly, young, fresh leaf samples were analyzed on BD FACSAria Fusion Sorter System (BD Biosciences Ltd., Franklin Lakes, NJ, USA) in the Cooperative Center for Research Facilities in the School of Medicine at Sungkyunkwan University. We followed the general procedures described in Doležel et al. (2007); each analysis was repeated three times. It was previously determined that three previously sampled populations of S. koreanus (Geumbyeong, Samak, and Ungil mountains) were diploid ( Lee et al., 2019). Flow cytometry in this study confirmed that the genome of S. koreanus was diploid when compared to the tetraploid genome of S. renifolius in Japan ( Fig. 2). Two additional populations of S. koreanus (Gangwon-do, Yonghwasan Mt. and Gyeonggi-do, Chungryungsan Mt.) had 2C (pg.) average values of 4.63 and 3.99, respectively. In contrast, the 2C value of S. renifolius had an average value of 7.08 for a population in Ibaragi Prefecture and 6.96 for a population in Iwate Prefecture. The flow cytometry results showed twice the 2C value in S. koreanus, suggesting a different ploidy level, as previously determined using karyotyping ( Iwatsubo and Otsuka, 2005; Lee et al., 2019).
Key to the species of Symplocarpus in eastern Asia
1. Inflorescence emerging before leaves in early spring; apex of spathe oriented horizontally.
2. Leaf blade 30–40 cm long; spathe 15–20 cm long; spadix 1.3–3.3 cm long ··········· S. renifolius 앉은부채 2. Leaf blade 6.9–23.5 cm long; spathe 6–11 cm long; spadix 0.9–1.7 cm long ··· S. koreanus한국앉은부채
1. Inflorescence emerging simultaneously with leaves in early spring or summer, or after leaves in summer; apex of spathe erect.
3. Inflorescence emerging after leaves in summer; leaf blade narrowly ovate to ovate-oblong; fruits ripening in spring of following year after flowering ······················· ········································· S. nipponicus 애기앉은부채 3. Inflorescence emerging simultaneously with leaves in early spring or summer; leaf blade cordate to widely ovate; fruits ripening in summer of same year as flowering.
4. Leaves 3–5, blade 20–40 cm wide; spadix ellipsoid; base of spathe above ground; flowering late June to early July; endemic to Japan ······ S. nabekuraensis 4. Leaves 10–40, blade 15–18 cm wide; spadix spherical; base of spathe partly below ground; flowering late March to early April; endemic to Russian Far East ····································· S. egorovii
ACKNOWLEDGMENTS
We thank Kanchi Gandhi of the Harvard University Herbaria for his help with the Latin scientific name. Special thanks goes to Elena A. Pimenova the Botanical Garden-Institute FEB RAS, Russia, for the English translation of the original description of S. egorovii. We are greatly indebted to Soyoung Lee for the line drawings of Symplocarpus koreanus. We also thank Ted Oh Kim, Petra Junes, Hee- Young Gil, Kyoung-In Heo, Sun Ae Song, Darwin Kim, Dhanna Kim, So-Yeon Lim, and Ji-Hyeon Jeon for their support and help in the field. An earlier version of the manuscript was greatly improved by the comments and suggestions from two anonymous reviewers. We are especially indebted to David Boufford of the Harvard University Herbaria for his constructive suggestions and careful editing of the manuscript. This study was supported by the National Research Foundation of Korea (NRF) and funded by the Ministry of Education (NRF-2013R1A1A2008659). We dedicate this paper to Dr. Sangtae Lee, who was professor and curator of Ha Eun Herbarium (SKK) at Sungkyunkwan University for nearly three decades (1981–2009) and contributed significantly to systematic studies of plants in Korea.
Fig. 1.
Symplocarpus koreanus. A. Habit. B. Details of roots. C. Young plant. D. Flowering habit. E, F. Spathe and spadix. G. Pistillate phase spadix with stipe. H. Staminate phase spadix with stipe. I. Flower with four stamens. J–L. Infructescence. M. Seeds. 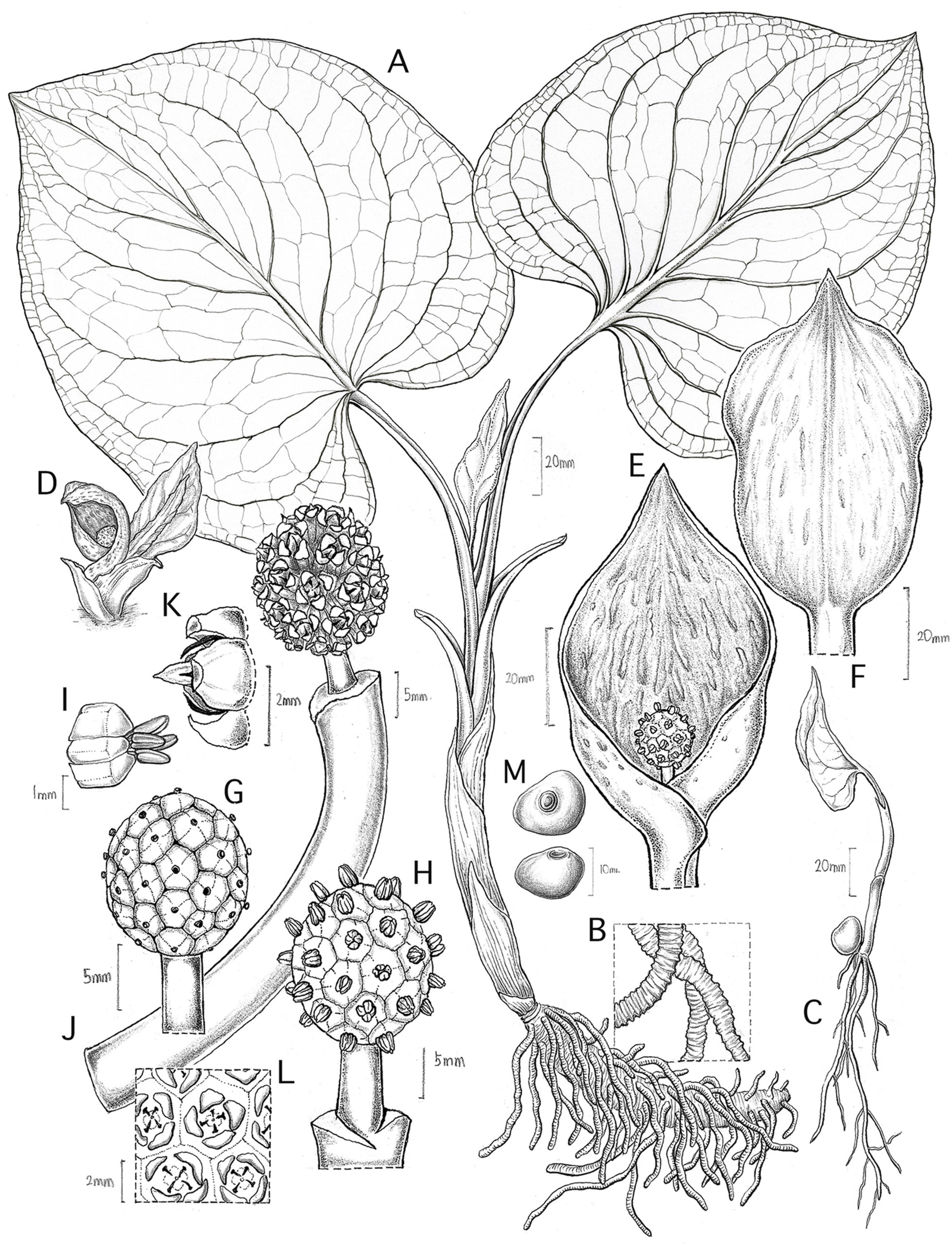
Fig. 2.
Flow cytometry histograms of Symplocarpus koreanus and S. renifolius. A. Solanum lycopersicum, internal standard (2C = 1.96 pg.). B. Symplocarpus koreanus (GW1; Gangwon-do population, Korea). C. S. koreanus (GG5; Gyeonggi-do population, Korea). D. S. renifolius (IBR5; Ibaragi population, Japan). E. S. renifolius (IWT6; Iwate population, Japan). 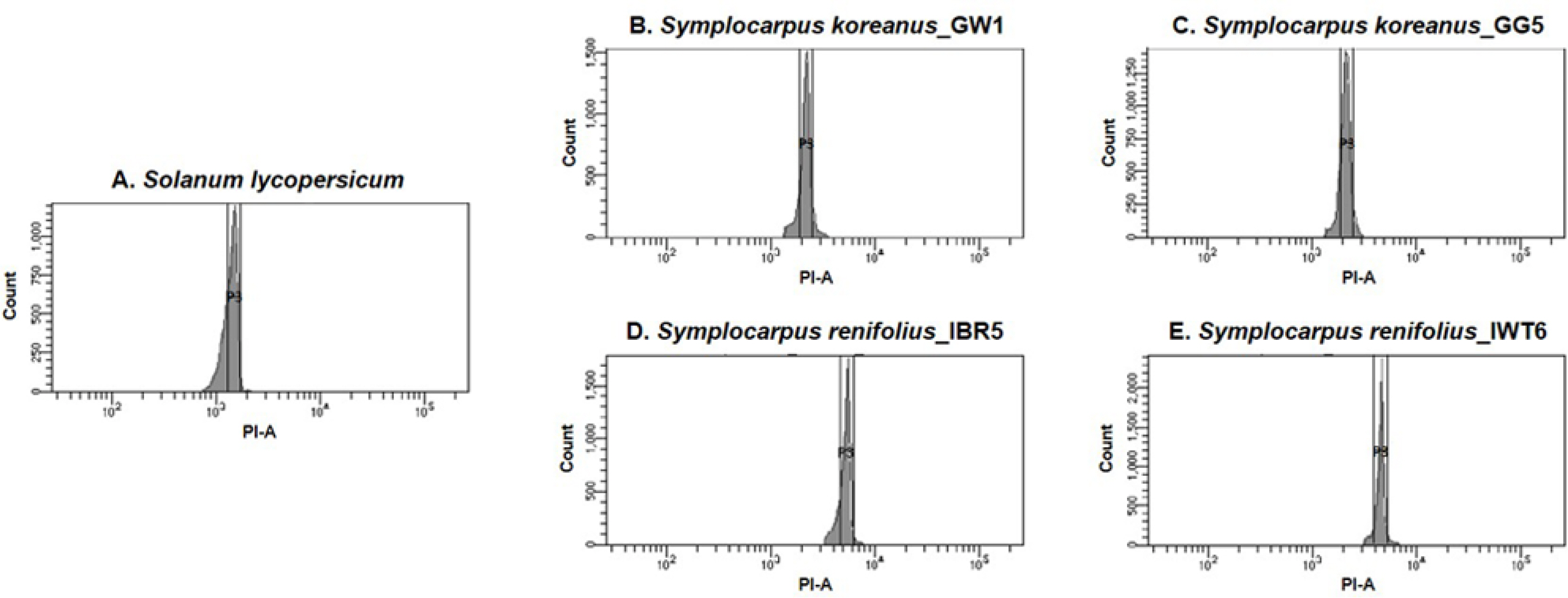
Table 1.
Comparison of key diagnostic features between Symplocarpus koreanus and S. renifolius
|
Character |
S. koreanus (mean ± SD) |
No. of plants |
S. renifolius (mean ± SD) |
No. of plants |
|
Root |
Root thickness (mm) |
1.56–5.26 (3.22 ± 0.73) |
6 |
- |
- |
|
Rhizome thickness (mm) |
9.90–26.99 (18.88 ± 6.82) |
|
|
|
|
Leaves |
Petiole length (cm) |
5.77–16.95 (11.15 ± 3.68) |
29 |
- |
- |
|
Petiole width (mm) |
1.88–10.20 (5.03 ± 1.87) |
|
|
|
|
Blade length (cm) |
6.93–23.47 (12.00 ± 4.03) |
29 |
- |
- |
|
Blade width (cm) |
4.56–18.79 (9.30 ± 3.63) |
|
|
|
|
Number of primary lateral veins |
5–7 (5.92 ± 0.64) |
13 |
- |
- |
|
Inflorescence and flowers |
Peduncle length (cm) |
3.46–20.79 (12.76 ± 5.34) |
18 |
- |
- |
|
Peduncle width (mm) |
0.33–2.04 (0.92 ± 0.33) |
|
|
|
|
Spathe length (cm) |
5.59–10.90 (7.50 ± 1.27) |
22 |
6.10–12.51 (8.85 ± 1.65) |
26 |
|
Spathe width (cm) |
2.79–5.82 (4.22 ± 0.74) |
|
3.83–7.59 (5.51 ± 1.06) |
|
|
Spadix length (mm) |
9.27–16.88 (12.21 ± 2.01) |
22 |
13.42–32.60 (21.59 ± 4.24) |
26 |
|
Spadix width (mm) |
7.44–15.04 (10.42 ± 1.76) |
|
9.52–23.07 (15.14 ± 2.86) |
|
|
Spadix ratio (length/width) |
1.02–1.44 (1.18 ± 0.09) |
|
1.14–1.70 (1.43 ± 0.14) |
|
|
Stipe length (mm) |
2.01–9.27 (5.66 ± 2.03) |
22 |
- |
- |
|
Stipe width (mm) |
2.04–4.36 (3.14 ± 0.58) |
|
|
|
|
Flower number/Spadix |
29–62 (40.04 ± 8.70) |
22 |
37–113 (79.34 ± 17.16) |
26 |
|
Flower size widest (mm) |
2.65–5.51 (3.84 ± 0.51) |
|
3.15–7.56 (4.72 ± 0.85) |
|
|
Flower size narrowest (mm) |
2.26–5.00 (3.37 ± 0.43) |
|
2.98–6.29 (4.22 ± 0.65) |
|
|
Chromosomes |
Chromosome number (2n) |
30 |
|
60 |
|
|
Ploidy level |
2x, diploid |
|
4x, tetraploid |
|
|
2C value (pg.) |
Gangwon population: 2.92–6.25 (4.63 ± 0.85) |
13 |
Ibaragi population: 6.11–7.63 (7.08 ± 0.39) |
12 |
|
Gyeonggi population: 2.72–6.02 (3.99 ± 0.68) |
16 |
Iwate population: 5.69–7.93 (6.96 ± 0.71) |
7 |
Table 2.
Comparison of major features of Symplocarpus koreanus and related species in eastern Asia.
|
Character |
S. koreanus
|
S. renifolius
|
S. egorovii
|
S. nipponicus
|
S. nabekuraensis
|
|
Distribution |
Korea |
China (Heilongjiang), Japan, Russia (Far East) |
Russia (South Primorian) |
China (Heilongjiang), Korea, Japan |
Japan (Nagano Pref., Honshu) |
|
Leaves: number |
2–4 |
Several |
10–40 |
2–5 |
3–5 |
|
Petiole: length (cm) |
5.8–17 |
Up to 40 |
30–50 |
Up to 30 |
20–50 |
|
Leaf blade shape |
Subcordate or cordate-ovate |
Orbicular-cordate |
Widely ovate |
Ovate to ovate-oblong |
Reniform–cordate |
|
Leaf base |
Cordate or reniform |
Deeply cordate |
Deeply cordate |
Obtuse to cordate-subcordate |
Cordate |
|
Blade length (cm) |
6.9–23.5 |
30–40 |
17–24 |
10–20 |
10–25 |
|
Blade width (cm) |
4.6–18.8 |
Up to 33 |
15–18 |
7–12 |
20–40 |
|
Primary lateral veins: number |
5–7 |
5 |
3–5 |
6 or 7 |
– |
|
Leaf emergence |
After emergence of spathe |
After emergence of spathe |
Simultaneous with spathe emergence |
Before emergence of spathe |
Simultaneous with spathe emergence |
|
Leaf senescence |
Summer |
Summer or Autumn |
Summer (mid-August) |
Summer |
Autumn |
|
Spathe: apex posture |
Elongated and pointing horizontally |
Elongated and pointing horizontally |
Short and pointing upward |
Short and pointing upward |
Short and pointing upward |
|
Spathe position |
Basal part in ground |
Basal part in ground |
Basal part in ground |
Basal part in ground |
Above ground |
|
Spathe length (cm) |
6–11 |
15–20 |
5–8 |
4–7 |
4–7 |
|
Peduncle length (cm) |
3.5–20.8 |
7–13 (–20) |
Up to 10 |
– |
4–8 |
|
Spadix shape |
Subglobose or ellipsoid |
Ellipsoid |
Spherical |
Ellipsoid |
Ellipsoid |
|
Spadix length (mm) |
9.3–16.9 |
13.4–32.6 |
25 |
– |
– |
|
Spadix width (mm) |
7.4–15 |
9.5–23.1 |
20 |
– |
– |
|
Flowering season |
Late Feb–early Mar |
Late Feb–early Mar |
Late Mar–early Apr |
Mid-Jul–late Aug |
Late Jun–early Jul |
|
Seed size: widest (mm) |
4.6–11.5 |
– |
10 |
– |
2–10 |
|
Fruiting |
Same year as flowering |
Same year as flowering |
Same year as flowering |
Following year after flowering |
Same year as flowering |
|
Spadix exothermic |
Yes |
Yes |
Unknown |
No |
Yes |
|
Chromosome number (2n) |
30 Lee et al. (2019)
|
60 Sokolovskaya and Probatova (1985), Iwatsubo and Otsuka (2005)
|
30 Marhold (2008)
|
(28), 30 Iwatsubo and Otsuka (2005), Jang and Weiss–Schneeweiss (2018)
|
60 Iwatsubo and Otsuka (2005)
|
Literature Cited
Doležel, J. Greilhuber, J and Suda, J. 2007. Estimation of nuclear DNA content in plants using flow cytometry. Nature Protocols 2: 2233-2244.   Ito, K. Ito, T. Onda, Y and Uemura, M. 2004. Temperature-triggered periodical thermogenic oscillations in skunk cabbage ( Symplocarpus foetidus). Plant and Cell Physiology 45: 257-264. Iwatsubo, Y and Otsuka, K. 2005. Chromosome numbers of Japanese Symplocarpus (Araceae). Journal of Phytogeography and Taxonomy 53: 203-205.
Jang, T.-S and Weiss-Schneeweiss, H. 2018. Chromosome numbers and polyploidy events in Korean non-commelinids monocots: a contribution to plant systematics. Korean Journal of Plant Taxonomy 48: 260-277. Knutson, RM. 1974. Heat production and temperature regulation in eastern skunk cabbage. Science 186: 746-747. Ko, SC. 2007. Araceae. The Genera of Vascular Plants of Korea. Flora of Korea Editorial Committee (ed.), Academy Publishing Co, Seoul. 1090-1094.
Kron, P. Suda, J and Husband, BC. 2007. Applications of flow cytometry to evolutionary and population biology. Annual Review of Ecology, Evolution, and Systematics 38: 847-876. Lee, JS. Kim, S-H. Lee, S. Maki, M. Otsuka, K. Kozhevnikov, AE. Kozhevnikova, ZV. Wen, J and Kim, S-C. 2019. New insights into the phylogeny and biogeography of subfamily Orontioideae (Araceae). Journal of Systematics and Evolution 57: 616-632. Li, H. Boyce, PC and Bogner, J. 2010. Symplocarpus. In Flora of China. Vol. 23. Acoraceae through Cyperaceae. Wu, ZY. Raven, PH. Hong, DY (eds.), Science Press, Beijing and Missouri Botanical Garden Press, St Louis, MO. 5-6.
Marhold, K. 2008. IAPT/IOPB chromosome data 5. Taxon 57: 553-562.
Mayo, SJ. Bogner, J and Boyce, PC. 1997. The Genera of Araceae. Royal Botanic Gardens/Kew Publishing, London. 822 pp.
Murata, J. 2016. Araceae. Flora of Japan, Vol. IVb. Angiospermae-Monocotyledoneae. Iwatsuki, K. Boufford, DE. Ohba, H (eds.), Kodansha, Tokyo. 6-7.
Nie, Z-L. Sun, H. Li, H and Wen, J. 2006. Intercontinental biogeography of subfamily Orontioideae ( Symplocarpus, Lysichiton, and Orontium) of Araceae in eastern Asia and North America. Molecular Phylogenetics and Evolution 40: 155-165. Ohwi, J. 1984. Araceae. Flora of Japan. Meyer, G. Walker, EH (eds.), Smithsonian Inst, Washington, D.C. 255-264.
Otsuka, K. Watanabe, R and Inoue, K. 2002. A new species of Symplocarpus (Araceae) from Nagano Prefecture, Central Japan. Journal of Japanese Botany 77: 96-100.
Otsuka, K. Hamada, T and Ueda, K. 2011. Thermogenesis in Japanese Symplocarpus species (Araceae). Journal of Japanese Botany 86: 224-229.
Pavlova, NS and Nechaev, VA. 2005. A new species of the genus Symplocarpus (Araceae) from the southern Russian Far East. Botanicheskii Zhurnal 90: 753-758.
Probatova, NS. Kozhevnikova, ZV. Kozhevnikova, AE and Rudyka, EG. 2012. Chromosome numbers of some vascular plant species from Amur River basin and Primorye (Russian Far East). Botanical Journal 97: 111-125 (in Russian).
Sokolovskaya, AP and Probatova, NS. 1985. Chromosome numbers in the Primorye territory, Kamchatka region, Amur valley and Sakhalin. Botanicheskii Zhurnal 70: 997-999.
Tsvelev, NN. 1996. Family Araceae. Vascular Plants of the Soviet Far East. 8: Kharkevich, SS (ed.), Nauka Press, Saint-Petersburg. 358-364 (in Russian).
Wada, N and Uemura, S. 1994. Seed dispersal and predation by small rodents on the herbaceous understory plant Symplocarpus renifolius. American Midland Naturalist 132: 320-327. Wada, N and Uemura, S. 2000. Size-dependent flowering behavior and heat production of a sequential hermaphrodite, Symplocarpus renifolius (Araceae). American Journal of Botany 87: 1489-1494.
APPENDICES
Appendix 1.
Specimens of S. renifolius examined in this study
JAPAN. Aomori Prefecture: Aomori City, Komagome, Michinokufukasawa Spa, 10 Jul 2004, K. Yonekura, TUS304706 (TU). Fukushima Prefecture: Omotego village, Kanayama, 3 May 1997, K. Yonekura et al., TUS188229 (TU). Hokkaido Prefecture: Atsuma village, Haronai Mt., 21 May 1985, H. Takahashi et al., TUS114928 (TU); Horokanai Town, Uryu Experimental Forest of Hokkaido University, 20 Jun 1996, K. Yonekura and E. Hayasaka TUS180566 (TU); Samani Town, Utoma, 13 May 1997, H. Yamaji, TUS390845 (TU). Hyogo Prefecture: Yoka Town, Myouken, 27 Apr 1986, N. Kurosaki, TUS125469 (TU). Ishikawa Prefecture: Komatsui City, Gohyaku Pass, 1 May 1994, K. Yonekura, TUS167165 (TU). Miyagi Prefecture: Marumori Town, Jiro-Taro Mt., 3 May 1984, T. Yokoyama s.n., TUS245294 (TU); Marumori Town, Touge, 30 Mar 2012, S. Y. Lee, TUS419857 (TU). Tochigi Prefecture: Imaichi City, Fubasami, 828 May 1982, H. Ohashi et al., TUS72187 (TU); Nakagawa Town, Umezo Park, 9 Apr 2012, S. Y. Lee, TUS419858 (TU).
|
|











