유전적 유사성으로 보아 멀구슬나무와 천련은 동일종
Melia toosendan and M. azadarach are a single species due to their genetic similarity
Article information
Abstract
한약재 천련자로 유통되는 중국 식물과 우리나라에도 분포하는 멀구슬나무의 연관성을 규명하기 위하여 중국남서부지역에서 천련자(Melia toosendan)로 채취하는 재료와 중국 남동부, 한국, 인도 등지에서 심고 있는 멀구슬나무(M. azadarach), 그리고 무환자나무과의 근연종들을 대상으로 핵 ITS, 엽록체 matK, rbcL, atpF-H, psbK-I, psbA-trnH 등 6개 마커의 염기서열을 비교분석하였다. 이들 염기서열 자료를 계통분석한 결과 한약재 천련자로 알려진 식물과 멀구슬나무는 동일종으로 판단된다. 이 두 종은 학자들에 따라서는 동일종 또는 이종으로 처리하던 것으로, 본 연구자들의 유전자 비교분석결과는 동일종으로 취급하는 것이 보다 타당함을 지지한다. 또한, 다양한 이명으로 세계 각지에서 기재된 멀구슬나무에 가장 가까운 종은 약재로 잘 알려진 님나무(Azadirachta indica)로, 이 종은 이전에 멀구슬나무속으로도 처리된 바 있다.
Trans Abstract
The nucleotide sequences of six markers, including nuclear ITS, chloroplast matK, rbcL, atpF-H, psbK-I and psbA-trnH, were analyzed for the plants known as Melia toosendan collected in Southwest China; M. azadarach planted in Southeast China, Korea and India; and species related to Sapindaceae in order to clarify the species boundary between M. toosendan and M. azadarach. The result of a phylogenetic analysis using the nuclear ITS and five chloroplast marker sequences determined that the plants known as M. toosendan and M. azadarach are the same species. These two species have been treated as a single species or as two different species depending on the researcher. The result of the present study supports the contention that the two species are the same. In addition, a sister species to M. azadarach registered in various countries with various basionyms is Azadirachta indica, a well-known medicinal plant. It has previously been classified as a member of the genus Melia.
멀구슬나무(Melia azadarch L.)는 멀구슬나무족(Melieae Harms), 멀구슬나무속(Melia L.)에 속하는 분류군으로서 인도, 동남아시아, 중국, 호주 등의 아열대에서 열대지방에 넓게 분포하며, 북미, 남미, 유럽 등지에도 조경수나 가로수, 약용식물로 식재하거나 귀화되어 자라는 것으로 알려져 있다(Pennington et al., 1981; Peng and Mabberley, 2008). 원래 멀구슬나무는 히말라야 일대의 재배 개체를 대상으로하여 기재되었다. 한반도에는 일본에서 도입되어 경남, 전남, 제주도 민가 주변에 식재되었고, 현재는 귀화되어 저지대의 숲가나 도로가 또는 황폐한 지역 등에 서식하는 것으로 보고되어 있다(Lee, 1996; Kim, 1998; Lee, 2006; Kim, 2007). 아열대 지방 및 온대 지방의 아시아 및 아프리카에 폭 넓게 분포하는 멀구슬나무는 지리적 혹은 개체 간 외부형태적인 변이 폭이 넓고, 지리적으로 거리가 먼 집단이나 개체 간에 많은 학명이 기재되어 왔다(Pennington and Styles, 1975; Mabberley et al., 1995; Peng and Mabberley, 2008).
멀구슬나무류의 열매는 한약재로 천련자(川子) 라고 부르며 오래전부터 습열을 제거하고 통증을 완화하며 기생충을 구제하는 효능이 있어 가슴통증과 산통 및 만성회충증으로 인한 복통을 치료하는데 사용해왔다(Kim, 2008). 천련자는 대한민국약전 외 한약(생약) 규격집에 천련(川, Melia toosendan Siebold & Zucc.) 또는 멀구슬나무(Melia azedarach L.)를 기원식물로 규정하고 있으며(Korea Food & Drug Administration, 2012), 중화인민공화국약전(Chinese Pharmacopoeia Commission, 2012) 부록에서는 천련을 기원식물로 규정하고 있다. 국내 생약시장에는 중국에서 수입하는 천련의 열매가 천련자로 유통되고 있으며 국내에서 생산되는 멀구슬나무의 열매 또한 천련자로 유통되고 있다. 천련(M. toosendan)은 중국에서 도입되어 일본에 재배하는 개체를 채집하여 Siebold and Zuccarini (1843)에 의해 처음 기재되었다. 천련은 멀구슬나무에 비하여 열매가 좀 더 크고, 소엽가장자리에 결각이 뚜렷한 멀구슬나무에 비해 결각이 뚜렷하지 않은 특징으로 두 종을 구별하여 왔으나, 최근 Peng and Mabberley (2008)에 의해 멀구슬나무(M. azadarach)의 이명으로 처리되었다. 따라서 천련자의 기원식물인 천련과 멀구슬나무의 동종성을 검증할 분자계통학적 연구가 요구된다. 한편, 멀구슬나무속이 포함되는 상위분류군에 대한 계통연구로는 Pennington and Styles (1975)이 멀구슬나무과 전반에 관하여 분류학적 유연관계를 정리한바있다. 또한, Muellner et al. (2003, 2008)은 멀구슬나무과와 멀구슬나무아과의 분자계통학적 연구를 수행하여 멀구슬나무속과 님나무속이 자매군을 이루며 멀구슬나무족에 위치하는 것을 보고하였으나, 멀구슬나무속에는 멀구슬나무 2개체만이 다루어져 종간이나 개체군 수준에서의 유연관계가 파악되지는 못하였다.
따라서, 본 연구에서는 한국, 중국 남부, 인도 지역에 분포하는 멀구슬나무 종집단과 천련 및 근연속, 근연과내 일부 분류군들을 대상으로 핵 DNA(nrDNA) ITS 지역과 엽록체 DNA(cpDNA)의 matK, rbcL, atpF-H, psbK-I 및 psbA-trnH 지역의 염기서열 분석을 수행하여 한약 재료인 천련자의 기원식물인 천련과 널리 재식하는 멀구슬나무의 동종성을 검토하고자 하였다.
재료 및 방법
재료
본 연구에서는 중국 운남성 지역에서 채집한 천련(M. toosendan) 8개체와 중국 남동부, 인도, 국내에서 채집한 멀구슬나무(M. azadarach) 8개체, 님나무(Neem, A. indica) 1개체, 참죽나무(Toona sinensis) 1개체, 외군으로서 멀구슬나무과와 유연관계가 깊은 것으로 알려진 무환자나무과의 무환자나무(Sapindus mukorossi) 1개체, 모감주나무(Koelreuteria paniculata) 1개체 등을 생체 재료로 사용하였다. 중국 운남성 지역에서 채집한 천련 8개체는 현지에서 천련자의 재료로 채취되는 자생하는 개체들로서, DNA분석을 위한 시료만 채집되었다. 그 외 채집된 개체들은 석엽표본을 제작하여 고려대학교 식물표본관(KUS)에 증거 표본으로 보관하였다(Table 1). DNA 분석은 nrDNA의 ITS 지역, cpDNA의 matK, rbcL, atpF-H, psbK-I, psbA-trnH 지역 등 총 6개 지역의 염기서열을 분석하였다(Table 2).
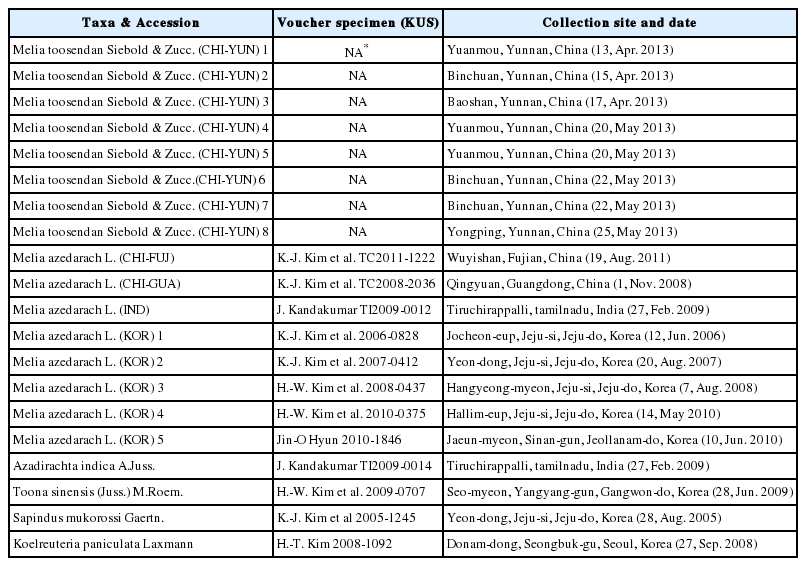
Materials and collection data used in this study.

GenBank accession numbers of sequences generated in this study.
DNA 추출, 증폭, 정제 및 염기서열 결정
생육지에서 채집한 잎을 엽병과 주맥을 제거시킨 후 -70℃에 냉동 보관하였다. 냉동 보관된 재료는 액체질소와 막자사발을 이용하여 분쇄한 후 CTAB 방법(Doyle and Doyle, 1987)과 G-spin™ IIp Genomic DNA Extraction Kit (Intron)을 사용하여 DNA를 추출하였다. 추출한 DNA는 모두 한국의 식물 DNA은행 (PDBK TC2008-2036, TC2011-1222, TC2013-1829-1836, TI2009-0012, TI2009-0014, 2005-1245, 2006-0828, 2007-0412, 2008-0437, 2008-1092, 2009-0707, 2010-0375, 2010-1846)에 보관하였다. PCR 증폭 마커로는 바코드 콘소시움에서 검증한 마커 중 cpDNA 5개 지역의 프라이머들을 사용하였고 (CBOL Plant Working Group, 2009), nrDNA의 ITS 지역의 경우에는 White et al. (1990)의 프라이머를 사용하였다. DNA 증폭을 위한 PCR은 94℃에서 5분 동안 initial denaturation시킨 후 94℃에서 denaturation 1분, 48−60℃에서 annealing 1분, 72℃에서 extension 1분을 주기로 하여 35회 반복한 후 72℃에서 7분간 extention의 조건으로 수행하였다. PCR 산물은 MEGA quick-spin™ Total Fragment DNA Purification Kit (Intron)을 이용하여 정제하였으며, 정제된 PCR 산물은 ABI PRISM 3730xl DNA sequencer로 염기서열을 결정하였다.
염기서열의 정렬 및 계통수 제작
양 방향으로 결정한 염기서열들은 Sequencher (v. 4.8; Gene Code Corporation, Ann Arbor, MI, USA) 프로그램을 사용하여 조합하였다. 완성된 염기서열정보는 유전자 또는 지역별로 NCBI GenbBank에 등록하였으며 각각 20개의 등록번호는 Table 2에 표시하였다. 염기서열 정보들은 지역별로 유합한 후, Geneious pro 5.5 (Drummond et al., 2011)내의 MUSCLE (Edgar, 2004)을 이용하여 정렬하였다. 계통수 제작은 PAUP* ver 4.0b10을 이용하여 Maximum parsimony (MP) 분석과 Neighbor joining (NJ) 분석, Maximum likelihood (ML) 분석을 수행하였다. MP 분석은 gap을 제5의 형질상태(new state)로 취급하여 heuristic searches 방법으로 분석하였으며 ‘TBR’ branch swapping, ‘collapse of zero branch lengths’, ‘MULPARS’ 및 ‘ACCTRAN’ option 등을 적용하였다. 모든 형질은 ‘equal weight’로 처리하였으며 Consistency index (CI), Retention index(RI), Rescaled consistency index (RC)을 산출하였다. MP 계통수의 지지도를 평가하기 위한 bootstrap 값은 위에 제시된 MP 분석 방법으로 1,000번 반복하여 산출하였다(Felsenstein, 1985). 6개 지역 자료의 염기변이(nucleotide divergence)는 Kimura’s two-parameter method 을 이용하여 계산하였고(Kimura, 1980), 이 값을 기초로 NJ tree를 산출하였다(Farris et al., 1996). ML 분석은 Modeltest 3.7 (Posada and Crandall, 1998)을 이용하여 nrDNA ITS 지역의 염기서열은 TrN+G 모델로, cpDNA 5개 지역의 유합한 염기서열은 K81uf+G 모델을 선정한 후 각각 염기들의 염기전이(transition)와 염기전환(transversion) 값을 적용하여 수행하였다. NJ 계통수와 ML 계통수의 작성된 각 분계도의 지지도를 평가하기 위한 bootstrap 값은 1,000번 반복을 통하여 산출하였다(Felsenstein, 1985). nrDNA와 cpDNA 염기서열 자료 간의 유사도를 평가하기 위하여 PAUP* ver 4.0b10을 이용하여 위상의존 제한분석(topology dependent constraint analysis)을 수행한 후 IM (Miyamoto index) 지수를 산출하였다(Kim and Jansen, 1994; Kim, 1996).
결 과
염기서열 분석
본 연구에서 얻어진 천련 8개체와 멀구슬나무 8개체에 대한 핵 리보솜 DNA의 ITS지역과 엽록체 DNA의 matK, rbcL, atpF-H, psbK-I, psbA-trnH 지역의 염기서열 분석결과를 Table 3에 정리하였다. Sanger sequencing 에서 각 염기는 5% noisy line으로 cut-off 하여 읽었으며, 이 수준에서 염기의 polymorphism은 관찰되지 않았다. 각 지역의 염기서열을 정렬한 결과, ITS는 748−749bp, atpF-H는 635−637bp, psbA-trnH는 541−561 bp 이었고, matK, rbcL, psbK-I는 843 bp, 703 bp, 392 bp로 모든 개체들의 염기서열의 길이가 동일하였다. ITS지역은 6개의 지역 중에서 가장 많은 13개의 사이트에서 염기치환변이가 일어났으며 CHI-YUN3, CHI-YUN6 2개체와 CHI-YUN7, CHI-YUN8 2개체는 각각 4개, 5개의 사이트가 계통학적으로 정보를 갖는 형질로 나타났다. psbA-trnH 지역의 경우 6개 지역 중 가장 많은 20개의 indel이 있었다. 6개 지역의 염기서열을 유합한 결과, 이들 염기서열의 총 3,893 bp로 정렬되었다. 정렬된 6개 지역의 유합 자료에서 indel의 수는 38개였으며, 29개 사이트에서 염기치환 변이가 일어났고 이 중 계통학적으로 정보를 갖는 형질 수는 20개로 나타났다.
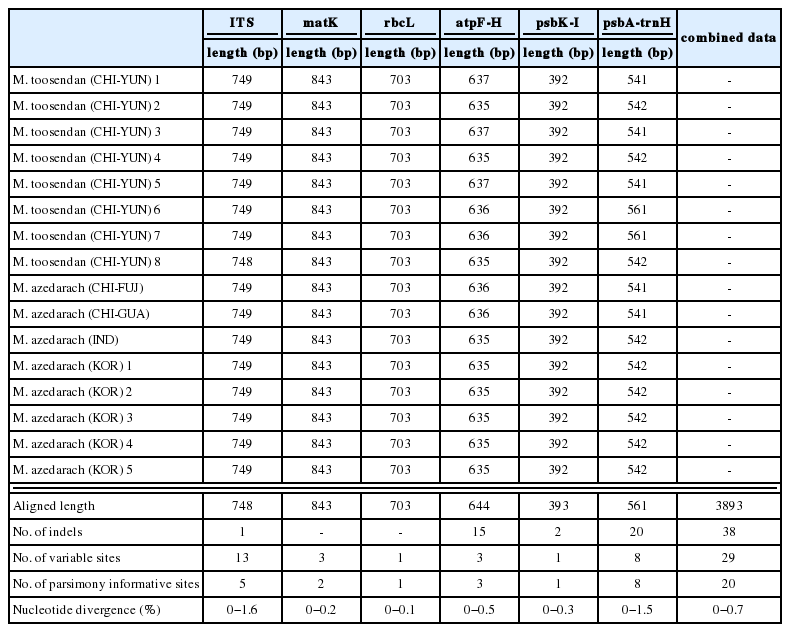
Statistical data for six markers (nrITS, matK, rbcL, atpF-H, psbK-I and psbA-trnH region) from sixteen taxa of Melia.
개체간의 염기변이를 Kimura’s two parameter method로 계산하여 백분율로 환산한 결과, 6개 지역을 유합한 멀구슬나무속 16개체의 염기변이는 0−0.7%이었으며(Table 3), 님나무, 참죽나무, 무환자나무 및 모감주나무를 포함한 20개체의 6개 지역을 유합한 멀구슬나무 16개체 염기변이는 0-0.45%이었다. 운남성에서 채집한 천련 1개체(CHI-YUN1)와 중국 남동부 지역에서 채집한 멀구슬나무 2개체(CHI-FUJ, CHI-GUA) 간의 염기서열 변이는 없었으며 국내에서 채집한 멀구슬나무 4개체(KOR1, KOR2, KOR4, KOR5) 간의 염기서열도 동일하였다. 멀구슬나무속 16개체군과 근연속 식물인 님나무 간의 염기변이는 3.09−3.15% 로 나타났다(Table 4).
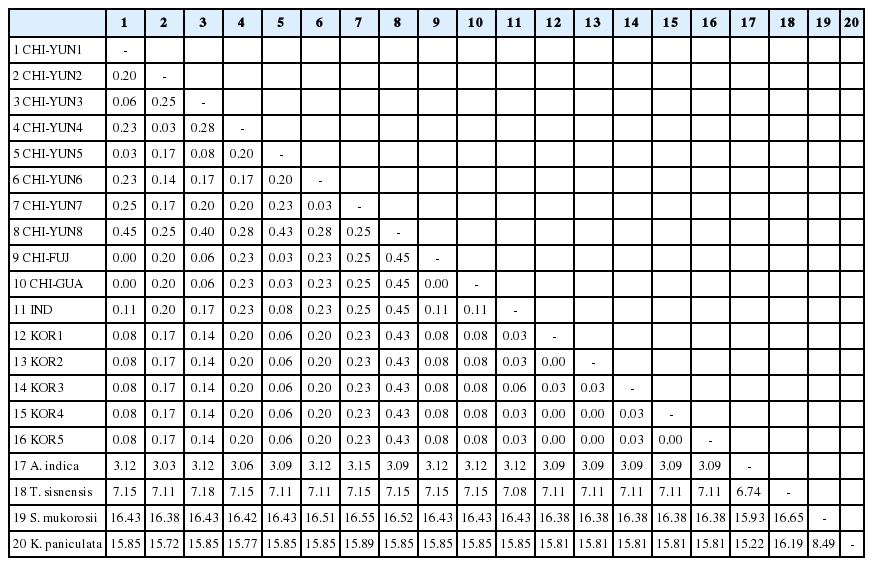
Nucleotide divergence of combined data (nrITS, matK, rbcL, atpF-H, psbK-I and psbA-trnH region) for twenty taxa (×100).
분자계통학적 분석
멀구슬나무속 16개체를 포함하여 멀구슬나무과 3종 18개체와 이들의 외군으로 무환자나무과 2종 2개체를 대상으로 nrDNA ITS 지역의 염기서열 자료와 cpDNA matK, rbcL, atpF-H, psbK-I, psbA-trnH 지역의 염기서열 유합자료를 기초로 하여 각각 ML, MP, NJ 계통수를 제작하였다. ITS 지역과 cpDNA 5개 마커를 유합한 지역들의 ML 분석결과 log likelihood (LN) 값은 각각 -2390.56203, -8159.62263이었다. ITS 지역의 ML 계통수에서 참죽나무는 100% bootstrap(BP) 값으로 가장 먼저 분기하였고, 이후 76% BP값으로 지지되는 님나무와 멀구슬나무속 집단으로 구성된 분계조로 분기하였다. 멀구슬나무속 집단은 단계통군을 형성하였으며 95% BP값에 의해 이들의 단계통성이 지지되었다. 천련 4개체(CHI-YUN1, CHI-YUN2, CHI-YUN4, CHI-YUN5)는 멀구슬나무 집단과 88% BP 값으로 유집되었으며, 나머지 천련 4개체는 52%의 낮은 BP값으로 분계조를 형성하였다(Fig. 1).
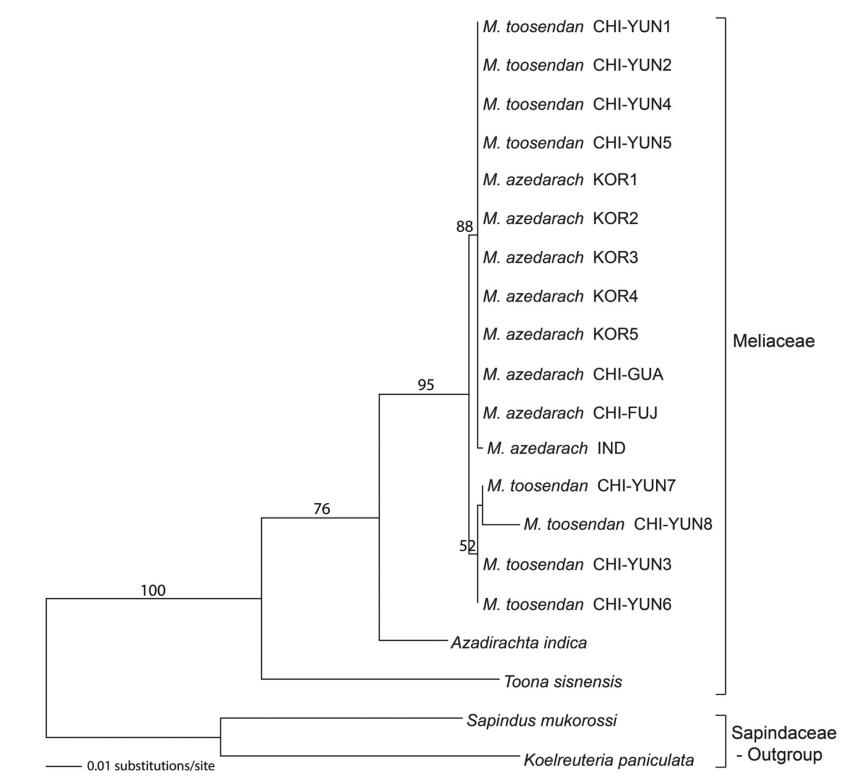
Maximum Likelihood (ML) tree of twenty taxa based on the nrITS region. The bootstrap values (>50%) are given above the branches.
CpDNA 지역들을 유합한 ML 계통수에서는 ITS 계통수와 동일하게 참죽나무와 님나무는 뚜렷이 구별되었으며, 멀구속나무속은 100%의 지지도로 단계통군을 형성하였다. 천련 2개체(CHI-YUN2, CHI-YUN4)는 ITS 계통수에서 멀구슬나무 집단과 분계조를 형성하였으나, ML 계통수에서는 운남성에서 채집한 천련 3개체(CHI-YUN6, CHI-YUN7, CHI-YUN8)와 53%의 낮은 BP값으로 분계조를 형성하였다. 천련 CHI-YUN3 개체는 ITS 계통수에서 천련 3개체(CHI-YUN6, CHI-YUN7, CHI-YUN8)와 유집되었으나, ML 계통수에서는 멀구슬나무 집단, 천련 2개체(CHI-YUN1, CHI-YUN5)와 95%의 높은 지지도로 하나의 분계조를 이루었다(Fig. 2). MP 분석 결과, ITS 지역은 443 단계로 구성된 4개의 MP 계통수를 얻었으며, CI, RI, RC 값은 각각 0.923, 0.821, 0.758 이었다. ITS 지역의 MP BP 분석 결과 얻어진 50% 이상 지지되는 node를 나타낸 계통수는 NJ 계통수와 같은 topology를 가지고 있었다. cpDNA를 유합한 지역의 MP 분석 결과, 1,296 단계로 구성된 10개의 MP 계통수를 얻었으며 CI, RI, RC 값은 각각 0.945, 0.916, 0.866 이었다. cpDNA 지역의 MP BP 분석 결과 얻어진 50% 이상 지지되는 node를 나타낸 계통수는 NJ 계통수와 동일하게 참죽나무와 님나무가 뚜렷이 구별되었으며 멀구슬나무속 집단은 100% 지지도로 2개의 분계조를 형성하였다(Fig. 2와 동일). ITS 지역의 MP, NJ 계통수와 cpDNA 지역들을 유합한 MP, NJ 계통수는 각 지역의 ML 계통수와 같은 topology을 보여 여기에 그림으로 제시하지는 않았다.
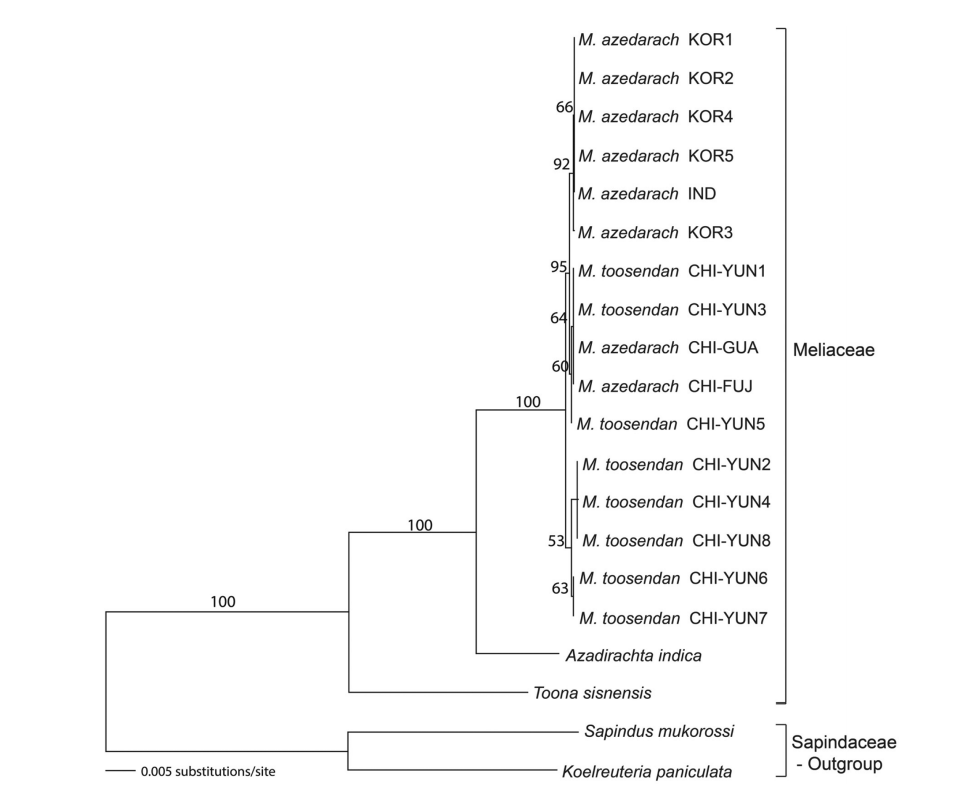
Maximum Likelihood (ML) tree of twenty taxa based on the five cpDNA regions (matK, rbcL, atpF-H, psbK-I, and psbA-trnH). The bootstrap values (>50%) are given above the branches.
nrDNA와 cpDNA 계통수 (Fig. 1 vs. Fig. 2) 간에 CHI-YUN3 개체를 포함 몇몇 개체의 위치가 달라서 두 염기서열 자료 간의 유사도를 평가하기 위하여 IM 지수를 계산하였다. IM 지수를 백분율로 환산한 결과 두 자료간에는 4.5% (32/704)로 불일치도(즉 95.5%의 일치도)를 나타냈다. 따라서 두 개의 자료를 독립분석하거나 유합분석하거나 큰 차이는 없는 것으로 평가되었다. 결과적으로 두 자료간의 차이는 거의 없으나 독립분석한 계통수간에 M. azedarach-M. toosendan 몇몇 개체의 위치가 바뀌는 것은 변이 형질이 적어 1−2개의 형질의 변이가 계통수에서 큰 차이를 만들기 때문이다. 유합분석할 경우 nrITS 계통수보다는 cpDNA 계통수에 가까운 topology가 얻어졌다.
고 찰
멀구슬나무속 집단과 근연속, 근연과내 분류군을 대상으로 ITS 지역 염기서열을 기초로 한 NJ, MP, ML 계통수와 cpDNA의 5개 지역을 유합한 염기서열을 기초로 한 NJ, MP, ML 계통수에서 멀구슬나무속 집단은 단계통군을 형성하였으며, 님나무가 각각 100% 값으로 자매군을 형성하였다(Figs. 1, 2). 이러한 결과는 멀구슬나무족내 멀구슬나무와 님나무가 자매군을 형성하는 Muellner et al. (2003, 2008)의 연구 결과와 일치한다.
멀구슬나무속 집단의 염기서열을 비교 분석한 결과, 중국남동부 지역에서 채집한 멀구슬나무 2개체(CHI-FUJ, CHI-GUA)와 천련 1개체(CHI-YUN1)는 동일한 염기서열을 공유하였으며 ML 계통수에서도 멀구슬나무집단에 함께 유집되었다. 또한 운남성에서 채집한 3개체(CHI-YUN1, CHI-YUN3, CHI-YUN5)와 동일한 지역에서 채집한 5개체간의 염기변이는 0.17−0.45% 로 한국 개체간의 염기변이 0.06−0.14% 보다 높게 나타나는 것으로 밝혀졌다 (Table 4). 그러나 천련과 멀구슬나무를 구별할 수 있는 사이트는 없었으며, 두 종 모두 단계통군을 형성하지 않고 서로 섞이는 결과를 보여주었다(Figs. 1, 2). 이러한 결과는 천련(M. toosendan)에서 관찰되는 큰 열매는 재배과정에서 나타나는 형질로 생각된다. 또한 잎의 결각도 멀구슬나무에서도 뚜렷한 것과 뚜렷하지 않는 중간형들이 있어서, 두 종을 구별하기에는 명확한 형질은 아니다. 그러나 두 종 모두 자생지가 없으므로 형태학적 형질도 분자형질들과 마찬가지로 지리적인 경향성의 확인은 어려울 것으로 보인다. 천련과 멀구슬나무 개체들을 대상으로 한 본 연구의 분자계통학적 연구 결과는 천련과 멀구슬나무는 서로 섞여 구분이 어렵고, 오히려 천련의 변이 폭에 멀구슬나무가 포함되는 양상이다. 아마도 이 결과는 멀구슬나무의 제한적인 시료채취에 기인한 것으로 생각된다. 본 연구 결과는 열매의 크기, 엽연의 형태 등에 있어서의 천련의 변이 폭과 멀구슬나무의 변이 폭이 중첩되므로 단일종으로 인식한 Peng and Mabberley (2008)의 견해를 지지한다. 나아가 본 연구 결과는 천련을 멀구슬나무의 이명으로 처리한 분류학적 견해를 지지한다.
한약재의 기원식물에 대한 정확한 종동정은 한약재의 품질 및 유통관리에 있어 중요한 과정이다. 최근 nrITS와 cpDNA와 같은 유전자 마커를 이용하여 한약재 기원식물의 종이나 원산지를 구별하는 연구들이 활발히 진행되고 있다(Moon et al., 2014; Son et al., 2014). 본 연구의 결과는 천련에서 기원한 천련자와 멀구슬나무의 열매가 동일한 종이라는 것을 분자계통학적으로 입증한 것으로 우리나라 약전과 중국 약전의 차이를 설명하는 근거자료가 될 것이다. 또한 멀구슬나무의 열매를 천련자로 이용하는데 합법적인 증거자료로 제시될 수 있다. 그러나 산지나 채취시기에 따라서 화학성분의 양적 및 질적인 차이는 있을 수 있으므로 이에 대한 분석이 같이 진행되는 것이 바람직할 것이다.
국내에서 채집한 멀구슬나무 5개체는 1개체(KOR3)가 matK 지역에서 1 bp 염기치환이 존재하고 나머지 지역에서는 동일한 염기서열을 갖는 것으로 나타나 본 연구에서 사용한 6개의 marker가 한국산과 중국산 멀구슬나무를 식별하는 데 유용한 marker가 될 수 있다는 것을 확인하였다. 이는 한국에 야생 또는 재배하는 멀구슬나무가 비교적 동일한 집단 또는 개체에서 기원하여 퍼져나갔을 가능성을 암시한다.
이상의 연구 결과를 종합하면 본 연구에 이용된 천련과 멀구슬나무 개체들은 단계통군을 형성하고, 유전적 다양성이 낮은 동일종으로 판단된다. 멀구슬나무(Melia azadarch L.)가 선취권이 있으므로 동일종으로 통합할 경우 이 학명을 이용하는 것이 옳다. 나아가 멀구슬나무의 자매군은 님나무로 판명되었다. 중국 운남에서 채집한 천련 3개체(CHI-YUN1, CHI-YUN3, CHI-YUN5)는 동일한 지역에서 채집한 다른 천련 개체들보다 한국에서 채집한 멀구슬나무 개체들과 더 낮은 염기변이를 나타내었다. 따라서 중국에서 천련자로 유통되는 천련(Melia toosendan)은 Peng and Mabberley (2008)의 견해에 따라 멀구슬나무의 이명으로 처리하는 것이 타당한 것으로 판단된다.
Acknowledgements
본 연구는 환경부 “차세대에코이노베이션기술개발사업(과제번호 416-111-007)”의 지원을 받아 수행되었습니다. 본 연구에 이용된 DNA는 한국의 식물 DNA은행(PDBK TC2008-2036, TC2011-1222, TC2013-1829--1836, TI2009-0012, TI2009-0014, 2005-1245, 2006-0828, 2007-0412, 2008-0437, 2008-1092, 2009-0707, 2010-0375, 2010-1846) 에 보존되고 있으며 이용 가능하다.