Achene wall anatomy and surface sculpturing of Launaea Cass. (Compositae: Cichorieae) with notes on their systematic significance
Article information
Abstract
This is the first study to deal in-depth with the achene ultra-structure of the genus Launaea Cass. in Egypt. It focuses on 12 taxa belonging to 10 species of this genus. The achene wall anatomy and surface sculpturing of those taxa were studied using scanning electron microscopy (SEM). Important aspects of the investigated taxa based on the achene characters as well as SEM micrographs of the achene surface and anatomical sections are given. Main and secondary costa of the inner achenes are used for easy differentiation between L. intybacea and L. massauensis
The taxonomy of the genus Launaea Cass. has always been problematic due to its confusing taxonomic history. This is reflected in the number of species that have been described. Launaea consists of 55 species and is mainly distributed in the S. Mediterranean area and in Africa and S.W. Asia (Kilian, 1997). In Egypt, Launaea is the largest genus (by 10 species) of the tribe Cichorieae (Zareh, 2005). Forsskål and Niebuhr (1775) recorded L. spinosa (Forssk.) Sch. Bip. ex Kuntze and L. mucronata (Forssk.) Muschl.; Boissier (1875) reported 10 taxa for the flora of Egypt, viz. L. spinosa, L. nudicaulis (L.) Hook. F., L. massauensis (Fresen.) Sch. Bip. ex Kuntze, L. procumbens (Roxb.) Ramayya & Rajagopal, L. capitate (Spreng.) Dandy, L. angustifolia subsp. arabica (Boiss.) N. Kilian, L. fragilis (Asso) Pau subsp. fragilis, L. fragilis (Asso) Pau subsp. tenuiloba (Boiss.) Zareh & Mohamed, L. mucronata (Forssk.) Muschl. and L. cassiniana (Jaub. & Spach) Kuntze. Muschler (1912) added L. fragilis and L. arabica (Boiss.) H. Lindb.. Amin (1957) in her thesis sited L. intybacea (Jacq.) Beauverd from Egypt, and finally Kilian (1997) reported L. amal-amine N. Kilian from Suez and Sinai. Recently, the genus has been subdivided in four sections, i.e., sect. Acanthosonchus (Sch. Bip.) O. Hoffm. (L. spinosa and L. acanthodes), sect. Microrhynchus (Less.) O. Hoffm. (L. nudicaulis, L. intybacea and L. massauensis), sect. Launaea (L. capitata) and sect. Zollikoferia (Pomel) O. Hoffm. (L. angustifolia, L. fragilis, L. mucronata and L. cassiniana) (Kilian, 1997).
Flowers in Launaea are yellowish with pubescent corolla tubes. Achenes brownish, blackish, or grayish; it may be homomorphic with 5 main ribs, accompanied by 2 secondary ribs (rarely undistinguished) or heteromorphic with inner achenes 4-main ribbed and marginal achenes 5-main ribbed. It also has pappus white, persistent or deciduous, homomorphic with setaceous rays or dimorphic with downy outer and setaceous inner rays.
Carpological characters enable most Cichorieae species to be identified even when no flowers are present (Blanca and Guardia, 1997). Other suitable applications include the identification of herbarium specimens in which the thickness of the peduncle may have been exaggerated by over-pressing, or specimens with an unusual number of bracts for the species of concern (Blanca and Guardia, 1997).
The term achene was defined by Wagenitz (1976) as a dry type of indehiscent monospermatous fruit derived from an inferior ovary. Achene features have been successfully used for the taxonomic classification of a wide range of Compositae groups (Lavialle, 1912; Briquet, 1916, 1930; Dittrich, 1968; Grau, 1980; Mouradian, 1995; Dittrich, 1996; Jan and Mukherjee, 2012; Zareh et al., 2016; Zareh, 1987). They have also been used for a number of different taxonomic levels for the tribe Lactuceae (Ownbey, 1950; Stebbins, 1953; Richardson, 1976; Tomb, 1977; De la Guardia and Blanca, 1987; Blanca and Guardia, 1997; Jan and Mukherjee, 2013).
In Launaea, the achenes morphology exhibits great diversity, and, as in the entire tribe, provides the most useful features for classification (Kilian, 1997). Appropriate descriptions of certain aspects of the achene morphology in Launaea, however, are rare in the literature, particularly concerning achene ribbing patterns. In the cross-sections of achenes, such deviations can be observed much more clearly (Kilian, 1997).
Anatomical sections of the achene were first employed by Lavialle (1912), who studied nearly 83 species of 15 genera of Compositae. Briquet (1916) studied the achene anatomy of the complexes Chrysanthemum and Anthemis. Achene anatomy studies have proved to be very rewarding to delimit the various taxa (Dittrich, 1968; Merxmuller and Grau, 1977; Haque and Godward, 1984; Mateu and Guemes, 1993; Abid and Qaiser, 2002; Zhu et al., 2006; Abid and Qaiser, 2008; Zareh, 1987). The most recent contribution to the knowledge on achenes anatomy in Launaea was provided by Kilian (1997), who studied the achene anatomies of representatives of all sections, only five species of which are represented in this study.
Studies on achene anatomies will be carried out to determine the strength of the systematic position of 12 taxa in the genus Launaea from Egypt. The major achene characteristics, including testa’s epidermis pattern, are described in detail.
Materials and Methods
The plants used in achene studies were L. spinosa, L. acanthodes (Boiss.) Kuntze, L. nudicaulis, L. intybacea (Jacq.) Beauverd, L. massauensis, L. capitata, L. angustifolia subsp. angustifolia, L. angustifolia subsp. arabica, L. fragilis subsp. fragilis, L. fragilis subsp. tenuiloba, L. mucronata and L. cassiniana (Table 1).
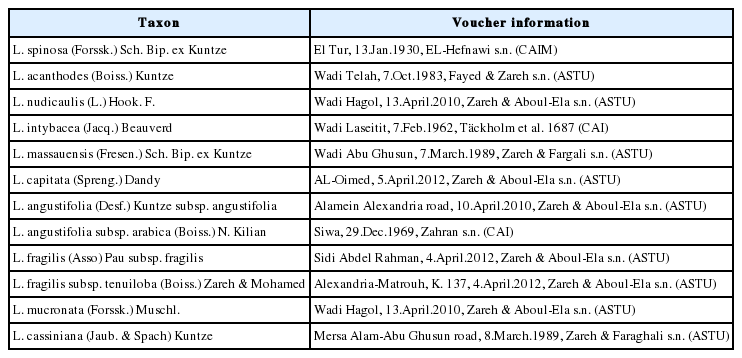
Voucher information for morphological examination of Launaea species in Egypt.
Dry achene materials for SEM were prepared by mounting them directly onto clean stubs using double-sided adhesive tape, followed by coating with gold in a JEOL JFC 1100E ion-sputtering device and then an examination using a JEOL JSM 5400 LV scanning electron microscope operated in an accelerated mode. The terms of Stearn (1966), Stant (1973), Barthlott (1981) and Boesewinkel and Bouman (1984) were used to describe the achene coat characteristics.
For the achene anatomy, dry achenes were boiled in water and then sectioned at 10–15 μ after being embedded in paraffin wax using a method described by Johansen (1940). The sections were stained using safranin (2% in 50% ethanol) and light green (1% in 95% ethanol). They then then permanently mounted in Canada Balsam.
The terminology of the achene wall anatomy essentially follows Lavialle (1912), Esau (1977), Kaul and Singh (1982) and Pak et al. (2001). The applied abbreviations to denote the details of the different layers of the achenes are as follows: e: epidermis, pm: primary mid rib, sm: secondary mid rib, par: parenchyma, scl: sclerenchyma, vb: vascular bundle, ec: endocarp, and sc: seed coat.
Results and Discussion
The achene of Launaea were brownish, blackish, or grayish; cylindrical to columnar; compressed or not, 2.5–8.0 × 0.4– 2.0 mm; subterete or prismatic; apex truncate, cuneate or attenuate; base truncate, tubular or horned; glabrous or papillose; homomorphic with 5 main ribs, accompanied by 2 secondary ribs (rarely undistinguished) or heteromorphic with inner achenes 4-main ribbed and marginal achenes 5-main ribbed, ribs smooth or transversely wrinkled; epidermal cells reticulate, bone-shaped or undifferentiated with straight raised or depressed anticlinal walls and concave periclinal walls, secondary cell wall rarely obscure in outlines with obtuse projections. Pappus was white, persistent or deciduous, homomorphic with setaceous rays or dimorphic with downy outer and setaceous inner rays. The morphological criteria of the achenes are summarized in Table 2.

External morphology of Launaea achenes.
Anatomically, pericarp was thick, laterally compressed; 4–5 stranded, each strand composed of a large main costa accompanied with 2 slightly smaller secondary costae; main and secondary costae separated and slightly or clearly differentiated in size; exocarp composed of a single epidermal layer; mesocarp of main and secondary costae with 2–25 layers of parenchyma and 3–18 layers of sclerenchyma cells, additional smallish groups of sclerenchymatous cells between main and secondary costae may present; each main costae is corresponding with a vascular bundle; endocarp composed of one layer of parenchyma. The anatomical criteria of the achenes are summarized in Table 3.

Anatomical features of Launaea achenes.
The mature achenes of Launaea exhibit great diversity and have proved to be the most useful characters for the distinction of some of the studied taxa. This diversity appears in the shape, size, colour, apex, base, epidermal cells and indumentum. The achenes of the studied taxa are commonly heteromorphic except for the two species of sect. Acanthosonchus (L. spinosa and L. acanthodes), which are subhomomorphic. The studied achenes are commonly ± cylindrical (oblong, oblanceolate or cuneate); only L. capitata and L. angustifolia exhibit winged achenes (Figs. 1, 2). Furthermore, the achene colour proved to be of a good taxonomic value, with most of the investigated marginal achenes being brown in colour, except for both L. capitata, which exhibits pale yellow achenes, and L. massauensis, with black achenes.

Scanning electron micrograph of marginal Launaea achenes. A. L. acanthodes. B. L. spinosa. C. L. nudicaulis. D. L. intybacea. E. L. massauensis. F. L. capitata. G. L. angustifolia subsp. angustifolia. H. L. angustifolia subsp. arabica. I. L. fragilis subsp. fragilis. J. L. fragilis subsp. tenuiloba. K. L. mucronata. L. L. cassiniana.
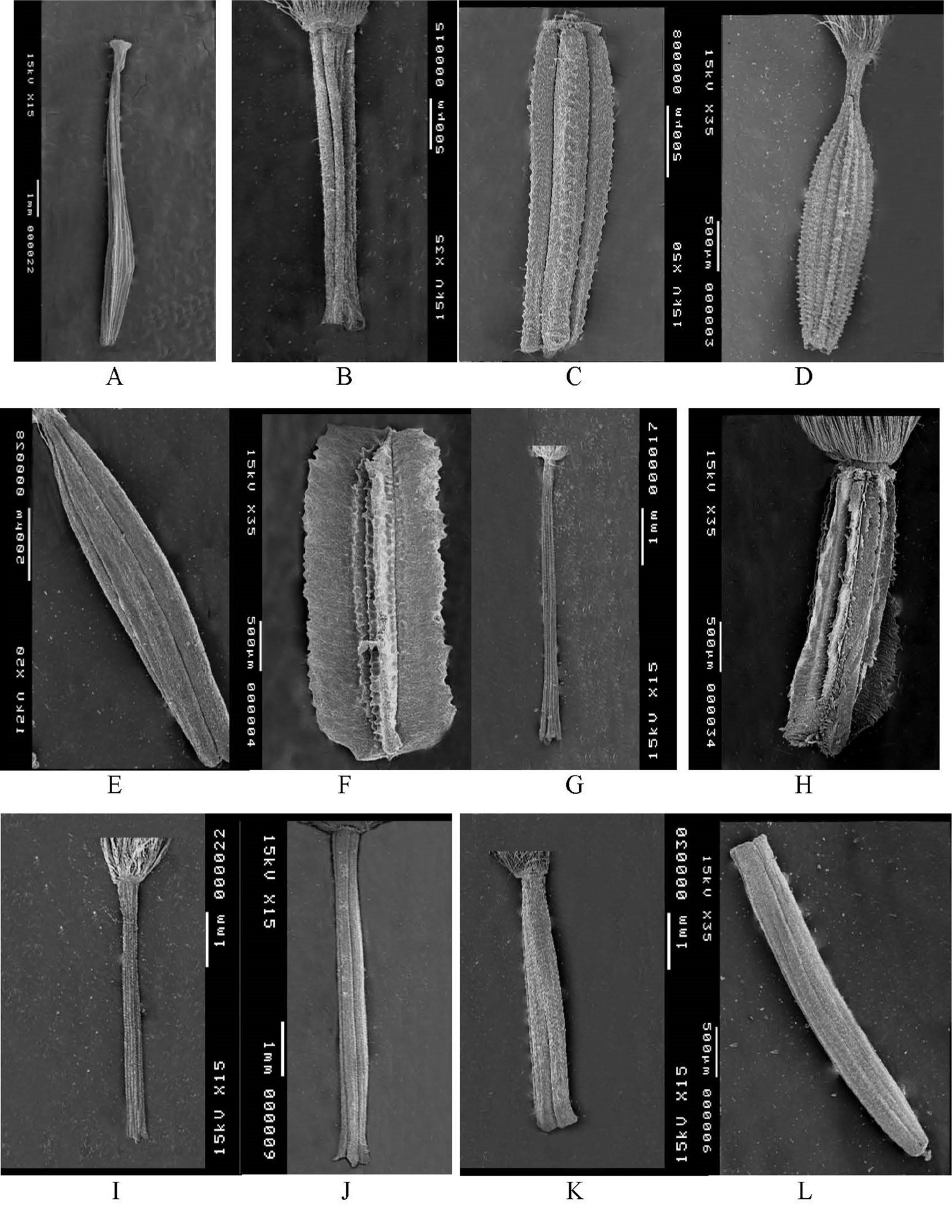
Scanning electron micrograph of inner Launaea achenes. A. L. acanthodes. B. L. spinosa. C. L. nudicaulis. D. L. intybacea. E. L. massauensis. F. L. capitata. G. L. angustifolia subsp. angustifolia. H. L. angustifolia subsp. arabica. I. L. frailis subsp. fragilis. J. L. fragilis subsp. tenuiloba. K. L. mucronata. L. L. cassiniana.
Most of the examined achenes have an approximate size of 2–6 × 0.5–1.3 mm (including the beak if present); in L. massauensis and L. procumbens, the achenes are apparently small, being 2–3 × 0.5–0.7 mm in size, while in L. acanthodes and L. capitata, the achenes are large at 4–8 × 0.9–1.3 mm.
The achene apex of the investigated taxa was found to be truncate, except for three species (L. acanthodes, L. intybacea and L. massauensis) which are characterized by an attenuate apex. In addition, the achene base proved to be tubular in two species (L. mucronata and L. cassiniana), horned in three species (L. spinosa, L. angustifolia and L. fragilis) and truncate in the remaining five species. Nevertheless, the achenes of the studied species are commonly wrinkled except for the species of sect. Zollikoferia (L. amal-aminae, L. angustifolia, L. fragilis, L. mucronata and L. cassiniana), which are characterized by papillose marginal achenes and wrinkled or smooth inner achenes (Figs. 3, 4).

Scanning elecron micrograph of marginal Launaea achenes surface. A. L. acanthodes. B. L. spinosa. C. L. nudicaulis. D. L. intybacea. E. L. massauensis. F. L. capitata. G. L. angustifolia subsp. angustifolia. H. L. angustifolia subsp. arabica. I. L. fragilis subsp. fragilis. J. L. fragilis subsp. tenuiloba. K. L. mucronata. L. L. cassiniana.

Scanning electron micrograph of inner Launaea achenes surface. A. L. acanthodes. B. L. spinosa. C. L. nudicaulis. D. L. intybacea. E. L. massauensis. F. L. capitata. G. L.angustifolia subsp. angustifolia. H. L. angustifolia subsp. arabica. I. L. fragilis subsp. fragilis. J. L. fragilis subsp. tenuiloba. K. L. mucronata. L. L. cassiniana.
Within the differentiated epidermal cells of marginal achenes, the anticlinal boundaries are generally undifferentiated in the species of sect. Zollikoferia (i.e., L. angustifolia, L. fragilis, L. mucronata and L. cassiniana), raised straight in L. spinosa, L. acanthodes, L. nudicaulis and L. capitata, and depressed straight in L. intybacea and L. massauensis, while the anticlinal boundaries of the inner achene are generally raised straight in all of the species except L. massauensis, being slightly sinuous margins. On the other hand, the periclinal walls are generally concave except for L. spinosa and L. acanthodes (Figs. 3, 4).
The pappus characters are much more useful for differentiation between some taxa; the setaceous type is only absent in L. angustifolia subsp. arabica, and the downy type is absent in four species (L. acanthodes, L. cassiniana, L. procumbens and L. capitata), with the pappus of the two later species having a pappus disk. Additionally, the pappus is persistent at the achene apex in all species except for L. procumbens, L. capitata and L. cassiniana, which are deciduous.
The present investigation showed that the anatomical features of Launaea achenes are of minor taxonomic value. The presence of additional groups of sclerenchyma cells between the main and secondary costae proved to be useful and reliable for the distinction between the sections. In the studied species of sect. Microrhynchus (L. nudicaulis, L. intybacea and L. massauensis) and sect. Launaea (L. capitata), they are often present, whereas they are absent in the species of sect. Acanthosonchus and Zollikoferia, according to the study of Kilian (1997).
Kilian (1997) studied the anatomical features of L. intybacea and did not study L. massauensis. It is easy to differentiate anatomically between the two species on the basis of the difference between the main and secondary costa of the inner achenes; the first is characterized by separated and clearly differentiated costa while the latter is not differentiated. Thus, we follow Täckholm (1974) with regard to L. intybacea as conspecific. Furthermore, the anatomical characters can be used easily to distinguish certain taxa (Figs. 5, 6). L. capitata is characterized by 2–3 large, distinct wrinkled wings, and L. angustifolia subsp. arabica has a papillate wing. Nevertheless, the pericarp thickness of the inner achenes can be used to differentiate between the much closer species of L. mucronata and L. cassiniana; the first is composed of 9–16 parenchyma and sclerenchyma layers while the latter is much thicker and composed of 17–26 layers (Figs. 5, 6).
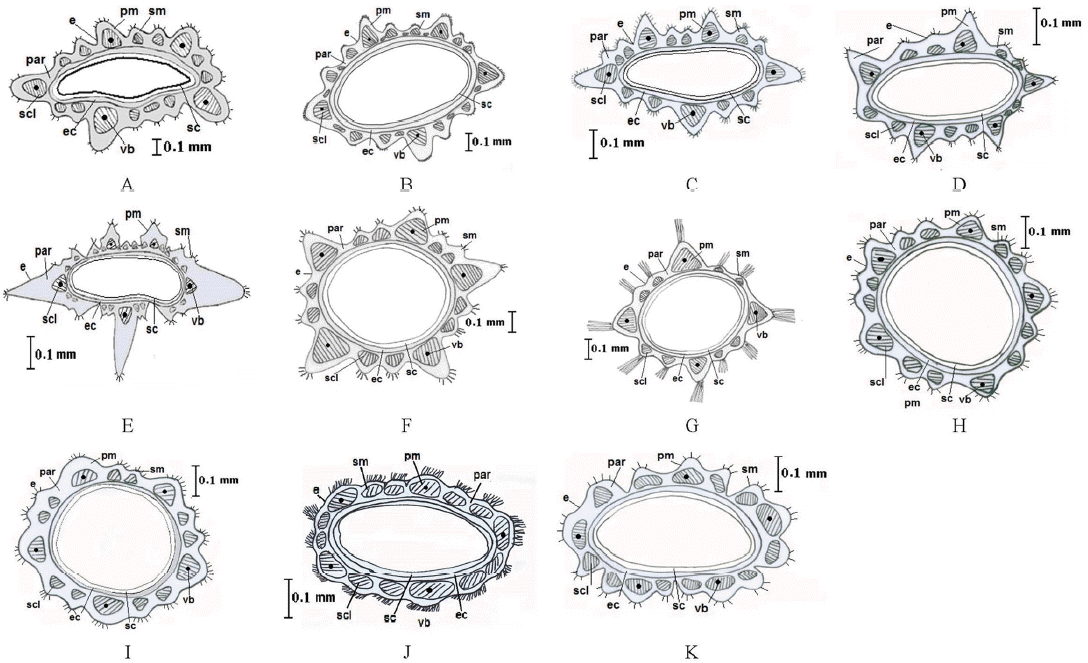
Diagrammatic transverse section of marginal Launaea achenes. A. L. spinosa. B. L. nudicaulis. C. L. intybacea. D. L. massauensis. E. L. capitata. F. L. angustifolia subsp. angustifolia. G. L. angustifolia subsp. arabica. H. L. fragilis subsp. fragilis. I. L. fragilis subsp. tenuiloba. J. L. mucronata. K. L. cassiniana. e: epidermis, pm: primary mid rib, sm: secondary mid rib, par: parenchyma, scl: sclerenchyma, vb: vascular bundle, ec: endocarp, sc: seed coat.

Diagrammatic transverse section of inner Launaea achenes. A. L. spinosa. B. L. nudicaulis. C. L. intybacea. D. L. massauensis. E. L. capitata. F. L. angusifolia subsp. angustifolia. G. L. angustifolia subsp. arabica. H. L. fragilis subsp. fragilis. I. L. fragilis subsp. tenuiloba. J. L. mucronata. K. L. cassiniana. e: epidermis, pm: primary mid rib, sm: secondary mid rib, par: parenchyma, scl: sclerenchyma, vb: vascular bundle, ec: endocarp, sc: seed coat.
Key to the taxa
1. Marginal achenes pale yellow, broadly 2–3-winged ················································································· L. capitata
1. Marginal achenes brownish to black, not winged (rarely 1-angeled) ········································································ 2
2. Marginal achenes surface wrinkled ························· 3
3. Achene subhomomorphic, main and secondary costae without additional groups of sclerenchyma cells between them ··············································· 4
4. Achene apex truncate, with obscure secondary cell wall and antrose obtuse projections; pappus persistent ············································ L. spinosa
4. Achene apex attenuate with fusiform secondary cell wall and triangular acute projections; pappus deciduous ······················· L. acanthodes
3. Achene heteromorphic, main and secondary costae with additional groups of sclerenchyma cells between them ························································ 5
5. Marginal achenes oblong, truncate, not beaked ························································ L. nudicaulis
5. Marginal achenes sub cylindrical, tapering or beaked ······························································· 6
6. Marginal achenes brown, epidermal cells of inner achenes bone-shaped with depressed straight anticlinal walls ··········· L. intybacea
6. Marginal achenes black, epidermal cells of inner achenes rectangular with raised straight to sinuous anticlinal walls ··· L. massauensis
2. Marginal achenes surface papillose ·························· 7
7. Achenes 1-angled, marginal achenes silky, hirsute at angles, base 5-notched and with 5 projections ···· 8
8. Inner achenes glabrous, marginal papillose; pappus dimorphic of setaceous and silky hairs ···················· L. angustifolia subsp. angustifolia
8. All achenes papillose; pappus monomorphic of silky hairs ········ L. angustifolia subsp. arabica
7. Achenes cylindrical, not silky or hirsute, base truncate ··································································· 9
9. Achenes longer than 6 mm long, innermost achenes 4-horned at base ······························ 10
10. Pappus as long as or slightly shorter than achene ················ L. fragilis subsp. fragilis
10. Pappus longer than the achene ·················· ························· L. fragilis subsp. tenuiloba
9. Achenes shorter than 5 mm long, all achenes not horned at base ················································ 11
11. Pappus persistent, dimorphic; inner achenes with pericarp of 9–16 layers ····································································· L. mucronata
11. Pappus deciduous, monomorphic; inner achenes with pericarp of 17–26 layers ····················································· L. cassiniana