쉬땅나무족(조팝나무아과: 장미과) 잎표피 미세형태학적 형질의 분류학적 유용성
Taxonomic significance of the leaf micromorphology in the tribe Sorbarieae (Spiraeoideae: Rosaceae)
Article information
Abstract
쉬땅나무족(Adenostoma: 3 spp., Chamaebatiaria: 1 sp., Sorbaria: 11 spp., Spiraeanthus: 1 sp.)과 연관 분류군인 Gillenia속 2분류군, Lyonothamnus속 2분류군을 포함한 총 20분류군의 잎 표피 미세구조에 대한 분류학적 유용성을 검토하고자 주사전자현미경(SEM)을 이용하여 관찰하고 기재하였다. Adenostoma속, Chamaebatiaria속, Spiraeanthus속에서 기공복합체(stomatal complex)는 향축면과 배축면 모두에 기공이 존재 하는 양면기공엽(amphistomatic type)이었고, Gillenia속, Lyonothamnus속과 Sorbaria속에서는 배축면(abaxial side)에만 존재하는 이면기공엽(hypostomatic type)으로 나타났다. 공변세포의 크기는 7.84−48.7 × 5.86−38.6 μm으로 속과 종마다 다소 차이를 보이는데, Sorbaria tomentosa var. tomentosa (7.84−11.8 × 6.84−10.5 μm)가 가장 작은 공변세포로 나타났고, Adenostoma fasciculatum var. obtusifolium (30.3−48.7 × 18.8−38.6 μm)에서 가장 크게 나타났다. 기공복합체의 형태는 대부분 불규칙형(anomocytic)이 우세하며, 불규칙형과 방사형(actinocytic)이 모두 나타나거나 드물게 다륜형(cyclocytic)이 나타나는 분류군이 확인되었다. 부세포의 수층벽(anticlinal wall)은 직선형과 곡선형이 동시에 관찰(straight/curved) 되거나 파상형(undulate) 또는 굴곡형(sinuate)이 관찰되었다. 연구된 분류군에서 나타나는 모용의 종류는 크게 4종류로 단세포단모(unicellular non-glandular trichome), 성상모(stellate), 선모(glandular trichome), 농포성 선모(pustular glandular trichome) 가 확인되었다. 연구된 분류군 가운데 일부 분류군의 표피세포에서 막성 판(membraneous platelets), 판모양(platelets)의 표피상납질(epicuticular wax)이 관찰되었다. 비록 잎 표피 미세형태학적 형질 내 본 족의 공 유파생형질을 찾을 수 없었지만, 모용의 다양성(특히, 성상모, 선모)과 표피상납질 유무의 분류학적 유용성을 확인하였다. 잎 표피 내 다양한 미세형태학적 형질은 외부형태학적 형질과 더불어 본 족을 이해하는데 보다 유용한 정보를 제공할 것이다.
Trans Abstract
A comparative study of leaf epidermal microstructures in the tribe Sorbarieae (Adenostoma: 3 spp., Chamaebatiaria: 1 sp., Sorbaria: 11 spp., Spiraeanthus: 1 sp.) including related genera Gillenia (2 spp.) and Lyonothamnus (2 spp.) was carried out using scanning electron microscopy (SEM) in order to evaluate their significance in taxonomy. The leaves of Adenostoma, Chamaebatiaria, and Spiraeanthus were amphistomatic, whereas Gillenia, Lyonothamnus, and Sorbaria were hypostomatic. The size range of the guard cells is 7.84−48.7 × 5.86−38.6 μm; the smallest one was found in Sorbaria tomentosa var. tomentosa (7.84−11.8 × 6.84−10.5 μm), while the largest measured example was Adenostoma fasciculatum var. obtusifolium (30.3−48.7 × 18.8−38.6 μm). Anomocytic stomata complex were the most frequent type (rarely cyclocytic), with usually both anomocytic and actinocytic types occurring in one leaf. On the surfaces, both the adaxial and abaxial anticlinal walls of the subsidiary cells vary (e.g., straight/curved, undulate, sinuate). Four types (unicellular non-glandular trichome, stellate, glandular trichome, pustular glandular trichome) of trichomes are found in the leaves. The epicuticular wax can be divided two types: membraneous platelets (Lyonothamnus) and platelets (Sorbaria arborea var. arborea, S. arborea var. subtomentosa, S. kirilowii, S. tomentosa var. tomentosa, Spiraeanthus schrenkianus). The trichome diversity (in particular, stellate, gland) and the existence of epicuticular wax may have taxonomic significance, although the leaf epidermal micromorphological characteristics do not provide synapomorphy in this tribe. These leaf micromorphological features are most likely better understood in the Sorbarieae when used in conjunction with external morphological characters.
잎 표피 형질에 대한 연구는 분류학적으로 매우 중요한 역할을 해왔으며, 많은 피자식물군 내에서 분류학적, 계통학적 유용성이 검증되었다(Wilkinson, 1979; Stace, 1984; Zhou et al., 1994; Yang and Lin, 2005; Zoric et al., 2009; Anil Kumar and Murugan, 2013; Deng et al., 2014). 특히, 잎 표피에서 관찰할 수 있는 형질 중, 기공의 형태, 빈도 및 모용의 종류와 분포 등은 다양한 분류군에서 중요한 형질로 연구되었으며(Ma et al., 2004; Moon et al., 2009; Adebowale et al., 2014; Cho et al., 2014), 잎, 줄기, 과실 표면에 나타나는 표피상납질(epicuticular waxes) 역시 분류학적으로 가치 있는 형질로서 연구되었다(Barthlott et al., 1998; Wissemann, 2000; Ensikat et al., 2006; Tomaszewski and Zielinski, 2014). 이러한 형질은 다양한 분류계급 중 족 수준에서 적용되어 족 내 공유파생형질 탐색이나 속간, 종간 식별형질 파악을 위해 시도되었다(Husain et al., 1990; Taia, 2004; De S-Haiad et al., 2009; Moon et al., 2009; Heo et al., 2013).
쉬땅나무족(Sorbarieae Rydb.)은 장미과(Rosaceae Juss.) 내의 조팝나무아과(Spiraeoideae Arn.)에 속하는 분류군으로, 최근 분자계통학적 연구를 기반으로 한 장미과 분류체계에서는 Adenostoma Hook. & Arn., Chamaebatiaria Maxim., Sorbaria (Ser.) A.Braun, Spiraeanthus Maxim. 4개 속을 포함하고 있다(Potter et al., 2007). Adenostoma속은 북미 서부에 2−3종, Chamaebatiaria속은 1종이 미국 서부에 분포하고, Sorbaria속은 4−9여종이 중앙아시아와 동아시아에 걸쳐 자생하며, Spiraeanthus속은 카자흐스탄 고유속으로 1종이 분포한다(Jepson, 1993; Gu and Alexander, 2003; Kalkman, 2004). 쉬땅나무족은 처음 Rydberg (1908)에 의해 다음의 세속(Chamaebatiaria, Schizonotus Lindl., Porteranthus Britton)을 포함하는 족으로 인식된 이래로 많은 학자들에 의해 족의 한계 설정이 변경되어 왔다. 특히, Potter et al. (2007)의 분류체계에 따르면, Schulze-Menz (1964)에 의해 쉬땅나무족 내 분류군으로 인식되어 온 Lyonothamnus A.Gray속이 조팝나무아과 내에서 정확한 족 내 분류학적 위치를 찾지 못하고 있는 것으로 나타났고, Rydberg (1908), Hutchinson (1964), Schulze-Menz (1964), Takhtajan (1997)에 의해 수과의 열매를 갖는 특징으로 독립된 Adenostomateae Rydb.족으로 인식되었던 Adenostoma속이 쉬땅나무족에 포함되어, 장미과 내 다른 분류군들과는 구별되는 독립된 족임을 지지하였다. 또한, Hutchinson (1964), Kalkman (2004)에 의해 Chamaebatiaria속, Sorbaria속, Spiraeanthus속과 함께 Gillenieae Maxim.족으로 인식된 Gillenia Moench속은 최근 상족(Supertribe)인 Pyrodae C.S. Campb., R.C. Evans, D.R. Morgan & T.A. Dickinson에 위치하였다.
쉬땅나무족(Sorbarieae) sensu Potter et al. (2007)에 속하는 4속의 잎, 소엽의 외부형태는 차이를 보이는데, Adenostoma속의 경우, 상록성인 단엽은 선형으로 침엽과 유사한 형태를 나타내고(Jepson, 1993), Chamaebatiaria속은 상록성의 2−3회 우상복엽으로 양치식물의 포자엽과 유사한 형태를 나타낸다(Kalkman, 2004). Sorbaria속은 일반적인 우상복엽의 형태를 보이며(Lee, 2007), Spiraeanthus속은 좁은 선형의 우상복엽으로 20−35개의 소엽으로 구성되는데, Chamaebatiaria속과 유사하게 전체적으로 양치엽과 비슷한 형태를 보인다(Komarov, 1939). 연관 분류군으로 Lyonothamnus속의 경우 잎의 이형화를 보이는데, 전연의 단엽 형태인 L. floribundus subsp. floribundus와 깊게 우상으로 갈라져 보이는 형태인 L. floribundus subsp. aspleniifolius으로 구분되며(Raven, 1963; Jepson, 1993; Kalkman, 2004), Gillenia속은 3출엽을 지닌다(Jepson, 1993). 최근에는 잎의 외부형태학적 형질 차이에서 파생되는 내부구조의 차이 파악 및 족 내 공유파생형질 탐색을 위해, 잎의 해부학적 형질에 대한 연구가 수행되었다(Song and Hong, 2014). 하지만, 잎의 미세형태학적 형질에 대한 연구는 아직 전무하다.
따라서, 본 연구의 목적은 쉬땅나무족 4속과 연관 분류군 2속(Gillenia, Lyonothamnus)을 대상으로 주사전자현미경(SEM)을 이용하여, 잎의 표피에서 확인할 수 있는 표피세포, 기공, 모용, 표피상납질 등의 잎 표피 미세구조를 상세히 관찰하고 기재하고, 이를 통해 미세형태의 중요성과 족 내 공유파생형질 탐색 및 속간, 종간 식별형질로서의 가치를 파악하는데 있다.
재료 및 방법
본 연구에 사용된 잎 재료는 Adenostoma속 2종 1변종, Chamaebatiaria속 1종, Gillenia 2종, Lyonothamnus속 1종, 1아종, Sorbaria속 6종, 4변종, 1품종, Spiraeanthus속 1종, 총 20분류군(36개 표본)으로 국내·외표본관(A, C, GH, K, KH, KHUS, MO, SNUA, abbreviations according to Thiers, 2015)으로부터 대여한 석엽표본에서 확보하였고, 일부 분류군들은 직접 야외 채집하여 FAA (40% formalin: 40% glacial acetic acid: 70% ethyl alcohol) 혼합액에 고정한 후, 70% 알코올에 장기 보관하면서 관찰하였다.
성숙한 잎을 선별하고, 모용과 선점을 관찰하기 위해 해부현미경(SM, SteREO Discovery. V8, Carl Zeiss Microscopy GmbH, Jena, Germany)을 사용하였고, 주사전자현미경(SEM)을 이용하여 잎 표피 미세형질들을 관찰하였다. 이미 건조된 표본이나, 대출표본으로부터의 재료는 약 90oC에서 3분간 끓이거나 연화제(Agepon, Agfa Gevaert, Leverkusen, Germany; Agepon wetting agent : distilled water, 1 : 200)에 넣어, 24시간 이상 담가 조직을 부드럽게 한 후, 70% 알코올에 보관하여 실험에 사용하였다. 사용된 재료는 Appendix에 제시하였다. 잎의 미세형태학적 형질을 관찰하기 위해서 70% 알코올에 보관된 시료는 아세톤시리즈(acetone series: 50%, 70%, 90%, 100%에서 각각 약 1시간 정도 보관)을 거쳐 탈수한 후, 이산화탄소(CO2) 가스를 이용한 임계점 건조 처리(Critical Point Drying; SPI-13200JAB, SPI Supplies, West Chester, USA)를 하여 완전 건조시켰다. 건조된 시료는 알루미늄 스터브(aluminum stub) 위에 올린 다음 이온 증착기(Ion-Sputter; E-1045, Hitachi, Tokyo, Japan)를 이용하여 금(Au)으로 2분간 이온증착(Ioncoating) 하고, 전계방출형 주사전자현미경(Field Emission Scanning Electron Microscope; FE-SEM, S-4700, Hitachi, Tokyo, Japan)으로 WD (working distance) 10−12 mm, 가속 전력 10 kv에서 관찰하였다.
기공의 크기와 모용의 길이는 Macnification program (2.0 version, Orbicule, Leuven, Belgium)으로 측정하였다. 일부 분류군을 제외하고는 지리적 변이를 고려하여 적어도 2개 이상의 표본에서 크기와 길이를 측정하였으며, 기공의 크기는 25개를 최소단위로 측정하여 최소−최대값을 나타냈고, 기공의 빈도는 일정 면적(0.05 mm2)에 분포하는 수를 10반복으로 측정하여 최소−최대값으로 표기하였다. 기공의 길이(L), 너비(W), 빈도(F) 간의 관계를 나타내기 위해 피어슨 상관관계(Pearson correlation)를 이용하여 분석하였다(IBM SPSS Statistics for Windows, Version 19.0., Released 2010, IBM Corp., Armonk, NY, USA).
잎 표피 미세형질에 관한 용어는 Roe (1971), Wilkinson (1979), Stace (1984)를 주로 따랐고, 기공의 빈도의 경우 Croxdale (2000)의 방법을 참고하였다.
결 과
본 연구결과 얻어진 쉬땅나무족 20분류군의 일반적인 잎 표피의 미세형태학적 특징들은 Table 1에 정리하였으며, Figs. 1−4에 제시하였다.
Overview of representative leaf characters, distribution and kinds of trichomes on leaves of all taxa studied in the tribe Sorbarieae and related genera.
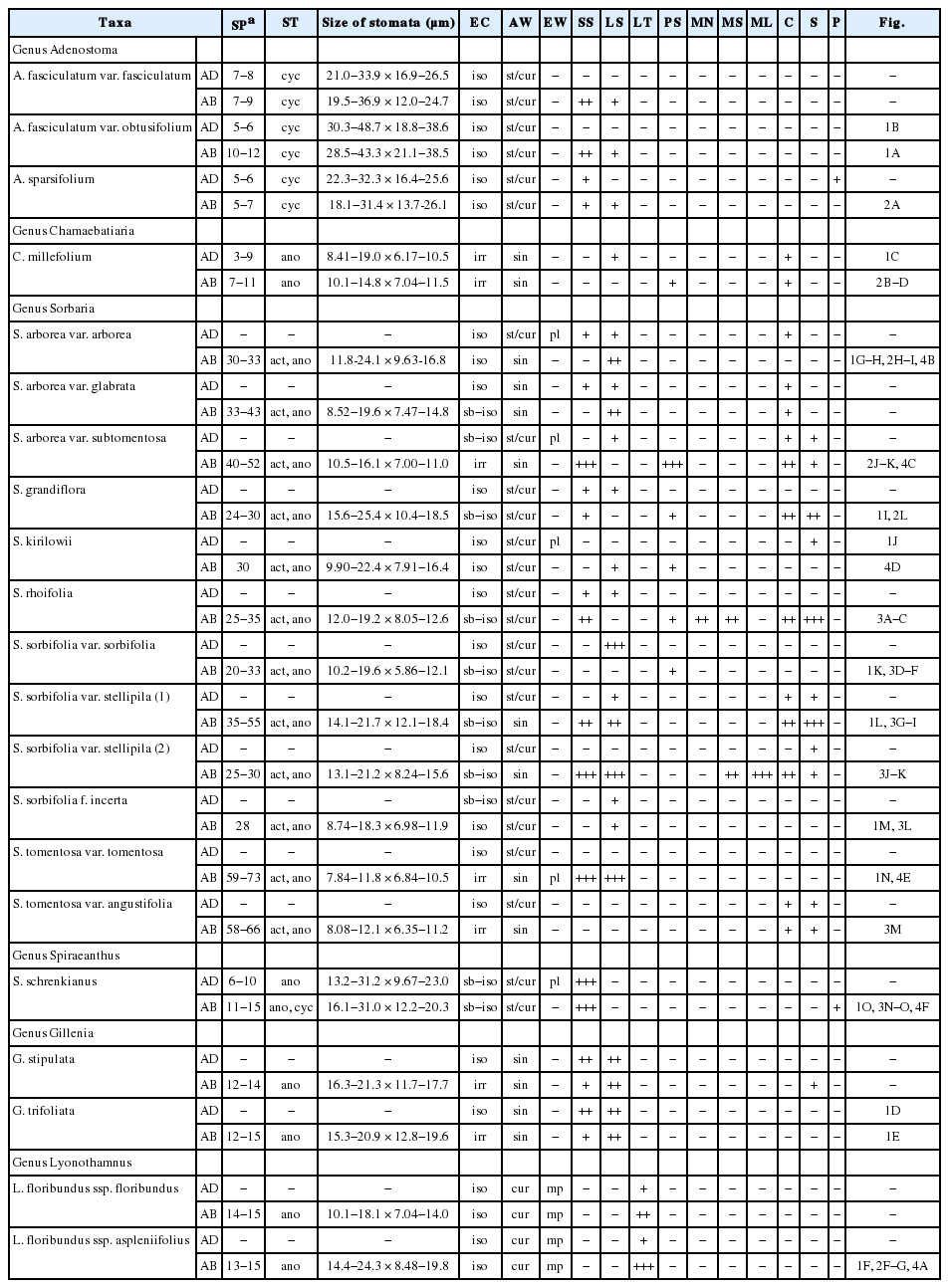
SEM micrographs of leaf surfaces of tribe Sorbarieae and related genera. A−B. Adenostoma fasciculatum var. obtusifolium. A. Abaxial surface showing isometric epidermal cells with cyclocytic stomata. B. Adaxial surface showing straight and curved isometric epidermal cells with cyclocytic stomata. C. Chamaebatiaria millefolium. Adaxial surface showing irregular epidermal cells with anomocytic stomata. D−E. Gillenia trifoliata. D. Adaxial surface showing isometric epidermal cells. E. Abaxial surface showing irregular epidermal cells with anomocytic stomata. F. Lyonothamnus floribundus ssp. aspleniifolius. Abaxial surface showing distribution of long twisted trichomes and stomata. G−H. Sorbaria arborea var. arborea. G. Abaxial surface showing isometric sinuate epidermal cells with actino/anomocytic stomata. H. Stoma with extended wings of striae. I. S. grandiflora. Abaxial surface showing sub-isometric epidermal cells with anomocytic stomata. J. S. kirilowii. Adaxial surface showing straight and curved isometric epidermal cells. K. S. sorbifolia var. sorbifolia. Stomata with extended wings of striae. L. S. sorbifolia var. stellipila (1). Abaxial surface showing sinuate anticlinal walls. M. S. sorbifolia f. incerta. Abaxial surface showing isometric epidermal cells with actinocytic stomata. N. S. tomentosa var. tomentosa. Abaxial surface showing irregular epidermal cells with anomocytic stomata having extended wings of striae. O. Spiraeanthus schrenkianus. Abaxial surface showing sub-isometric epidermal cells with cyclocytic stomata and short non-glandular trichome.

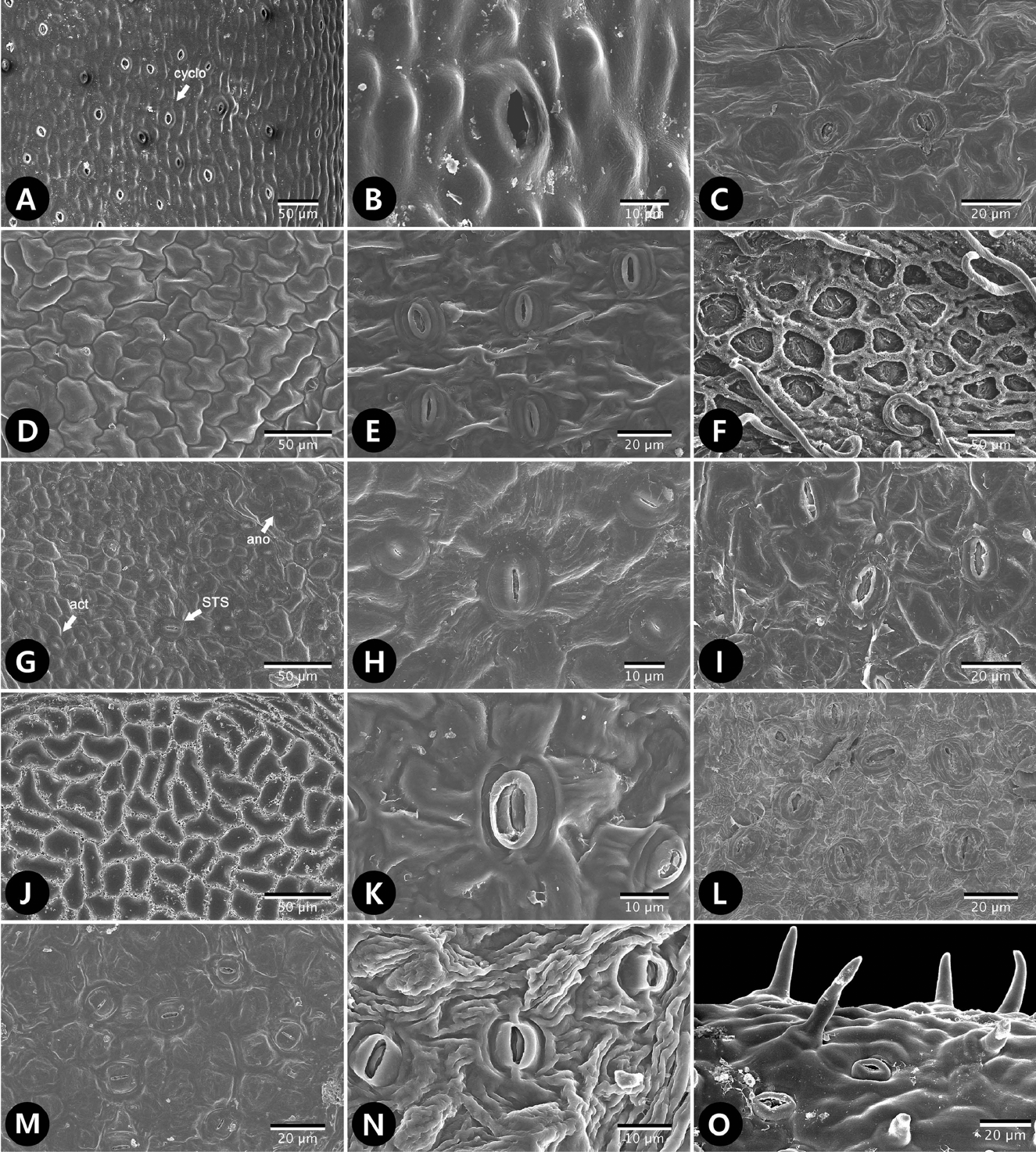
SEM micrographs of trichomes of tribe Sorbarieae and related genera. A. Adenostoma sparsifolium. Short trichome (< 100 μm) with smooth wall (SS, type 1-1) and pustular gland (P, type 4). B−D. Chamaebatiaria millefolium. B. Sessile porrect-stellate trichome (type 2-1). C. Capitate gland (type 3-1). D. Long trichome (> 100 μm) with smooth wall (type 1-2). E. Gillenia trifoliata. Long trichome at the mid vein. F−G. Lyonothamnus floribundus ssp. aspleniifolius. F. Long twisted trichome (> 100 μm) with spiral wall (type 1-3). G. Trichome surface showing spiral striae. H−I. Sorbaria arborea var. arborea. H. Short and long trichome with smooth wall at adaxial surface. I. Trichome surface showing smooth wall. J−K. S. arborea var. subtomentosa. J. Distribution of sessile porrect-stellate trichome (PS, type 2-1) and subsessile gland (S, type 3-2) at abaxial surface. K. Central part of sessile porrect-stellate trichome. L. S. grandiflora. Subsessile gland.
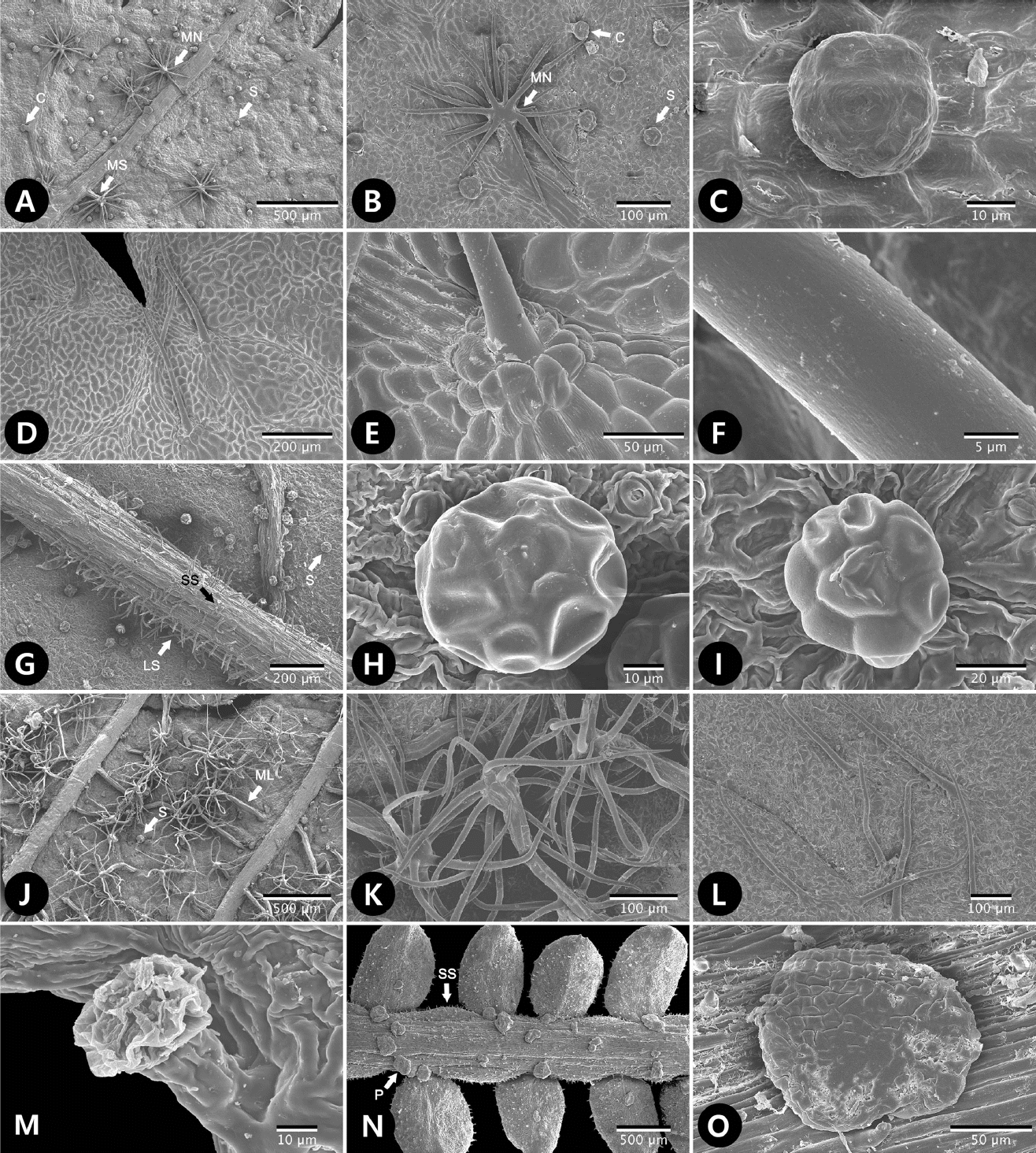
SEM micrographs of trichomes of tribe Sorbarieae and related genera. A−C. Sorbaria rhoifolia. A−B. Distribution of capitate gland (C, type 3-1), sessile multiangular stellate trichome without stalk (MN, type 2-2), multiangular stellate trichome with short stalk (MS, type 2-3) and subsessile gland (S, type 3-2) at abaxial surface. C. Subsessile gland. D−F. S. sorbifolia var. sorbifolia. D. Long trichomes (> 100 μm) with smooth wall (type 1-2) at adaxial surface. E. Subsidiary cells surrounding the non-glandular long trichome. F. Trichome surface showing smooth wall. G−I. S. sorbifolia var. stellipila (1). G. Distribution of long trichomes (LS, type 1-2) with smooth wall, subsessile gland and short trichome with smooth wall (SS, type 1-1) at abaxial surface. H−I. Subsessile gland. J−K. S. sorbifolia var. stellipila (2). J. Distribution of multiangular stellate trichome with long stalk (type 2-4) and subsessile gland at abaxial surface. K. Multiangular stellate with long, multiseriate stalk. L. S. sorbifolia f. incerta. Long trichomes with smooth wall. M. S. tomentosa var. angustifolia. Capitate gland. N−O. Spiraeanthus schrenkianus. N. Distribution of pustular gland (P, type 4) and short trichome with smooth wall (SS, type 1-1). O. Pustular gland.

SEM micrographs of epicuticular wax deposits of tribe Sorbarieae and related genera. A. Lyonothamnus floribundus ssp. aspleniifolius. Membraneous platelets. B−F. Platelets. B. Sorbaria arborea var. arborea. C. S. arborea var. subtomentosa. D. S. kirilowii. E. S. tomentosa var. tomentosa. F. Spiraeanthus schrenkianus.
표피세포
표피세포의 형태는 등방형(isometric, Fig. 1D), 아등방형(sub-isometric, Fig. 1I), 불규칙형(irregular, Fig. 1C)으로 관찰되었으며, 대부분의 분류군에서 향축면(AD: adaxial side)과 배축면(AB: abaxial side)에서 모두 유사하게 나타났다(Table 1). 다만, Gillenia속과 Sorbaria속에서는 향축면과 배축면의 표피세포 형태가 다소 다르게 관찰되었다. 부세포의 수층벽(AW: anticlinal wall)은 직선형과 곡선형이 동시에 관찰(straight/curved) 되거나(Fig. 1J), 파상형(undulate) 또는 굴곡형(sinuate)이 나타났다(Fig. 1D; Table 1).
기공복합체
Adenostoma속, Chamaebatiaria속, Spiraeanthus속에서 기공복합체(stomatal complex)는 향축면과 배축면 모두에 기공이 존재하는 양면기공엽(amphistomatic type)이었고, Gillenia속, Lyonothamnus속과 Sorbaria속에서는 배축면(abaxial side)에만 존재하는 이면기공엽(hypostomatic type)으로 나타났다(Table 1). 공변세포의 크기는 7.84−48.7 × 5.86−38.6 μm으로 속과 종마다 다소 차이를 보이는데, 가장 작은 공변세포는 Sorbaria tomentosa var. tomentosa에서 7.84−11.8 × 6.84−10.5 μm로 나타났고, 반면 가장 큰 공변세포는 Adenostoma fasciculatum var. obtusifolium로 크기가 30.3−48.7 × 18.8−38.6 μm로 나타났다(Table 1). 기공의 길이(L)와 너비(W)는 서로 양의 상관관계를 나타냈고(L과 W: r = 0.97, P < 0.01), 기공의 길이와 빈도(F)는 음의 상관관계를 나타냈다(L과 F: r = -0.58, P < 0.01; W와 F: r = -0.53, P < 0.01). 기공복합체의 형태로 Adenostoma속에서는 다륜형(cyclocytic)이 관찰되었고(Fig. 1A−B), Chamaebatiaria속, Gillenia속, Lyonothamnus속에서는 불규칙형(anomocytic)이 나타났다(Fig. 1C, 1E, 1F). 반면, Sorbaria속에서는 불규칙형과 방사형(actinocytic)이 동시에 나타났고(Fig. 1G−H), Spiraeanthus속에서는 대부분 불규칙형이 나타나고 드물게 다륜형이 확인되었다(Fig. 1O).
모용
연구된 20분류군의 잎 표피에는 향축면과 배축면에 다양한 종류의 모용이 분포하는 것으로 밝혀졌다. 비선모 모용은 단세포 단모(simple unicellular non-glandular trichome)와 성상모(stellate)가 나타났고, 선모는 짧은 자루의 두상선모(capitate glandular trichome)와 자루가 없는 두상선모(subsessile glandular trichome)가 나타났다. 모용의 종류에 따라 다음의 타입으로 구별하였다.
Type 1. 단세포단모(Simple unbranched unicellular non-glandular trichome)
단세포성 단모로 모용의 표면과 길이에 따라 3가지 서브타입으로 구별된다.
subtype 1-1. 평활상의 짧은 단모(short trichome with smooth wall): 길이 100 μm 이하의 짧은 단모로 모용의 표면이 매끈하다. Adenostoma속, Gillenia속과 Sorbaria속 일부 분류군에서 확인되었고(Figs. 2A, 2H−I, 3G), Spiraeanthus속에서는 향축면, 배축면 모두에서 높은 빈도로 관찰되었다(Fig. 3N). 한편, Chamaebatiaria속과 Lyonothamnus속에서는 관찰되지 않아 구별되었다.
subtype 1-2. 평활상의 긴 단모(long trichome with smooth wall): 길이 100 μm 이상의 길이를 지닌 단모로 모용의 표면이 매끈하다. Gillenia속에서 향축면, 배축면 모두에서 많은 수가 관찰되었고(Fig. 2E), Adenostoma속에서는 배축면에서, Chamaebatiaria속에서는 향축면에서 관찰되었다(Fig. 2D). Sorbaria속에서는 S. sorbifolia var. sorbifolia (Fig. 3D−F), S. sorbifolia var. stellipila (Fig. 3G), S. tomentosa var. tomentosa의 배축면에서 높은 빈도로 확인되어 S. arborea complex, S. kirilowii, S. sorbifolia f. incerta (Fig. 3L) 등의 속 내 다른 분류군과 구별되었다. Lyonothamnus속과 Spiraeanthus속에서는 나타나지 않았다.
subtype 1-3. 유선상으로 꼬인 긴 단모(long twisted trichome with striate wall): 길이 100 μm 이상의 길이를 지닌 단모로 한 쪽 방향으로 꼬인 형태로 모용의 표면이 유선상으로 관찰된다(Fig. 2F−G). Lyonothamnus속에서만 관찰되어 연구된 나머지 분류군과 구별되었다. 특히, 향축면보다 배축면에서 높은 빈도로 관찰되었다.
Type 2. 성상모(stellate)
별 모양의 비선모로 성상모를 이루는 단모(ray)의 빈도와 자루(stalk)의 유무로 4가지 타입으로 구별된다.
subtype 2-1. 수평으로 납작한 형태의 성상모(sessile porrectstellate trichome): 자루가 없는 성상모로 중앙에 위치한 단모(central ray)를 중심으로 수평으로 위치한 6−8개의 단모(horizontal rays)로 구성된다. Chamaebatiaria속의 배축면에서 관찰되고(Fig. 2B), Sorbaria속 내에서는 S. arborea var. subtomentosa (Fig. 2J−K), S. grandiflora, S. rhoifolia, S. sorbifolia var. sorbifolia의 배축면에서만 관찰되어 구별되었다.
subtype 2-2. 자루가 없는 다각상 성상모(sessile multiangulate trichome): 자루가 없는 성상모로 10개 이상의 단모(ray)가 다각상으로 뻗어 위치한다. 연구된 분류군 내에서 S. rhoifolia 에서만 관찰되어 구별되었다(Fig. 3A−B).
subtype 2-3. 짧은 자루의 다각상 성상모(multi-angulate trichome with short stalk, < 300 μm): 300 μm 이하의 비교적 짧은 길이의 자루를 가진 다각상 성상모로, 연구된 분류군 내에서 S. rhoifolia (Fig. 3A), S. sorbifolia var. stellipila (2) 의 배축면에서만 다수가 다른 분류군과 구별되었다.
subtype 2-4. 긴 자루의 다각상 성상모(multi-angulate trichome with long stalk, > 300 μm): 300 μm 이상의 긴 자루를 가진 다각상 성상모가 배축면의 측맥을 따라 분포하는데, S. sorbifolia var. stellipila (2) 에서만 높은 빈도로 관찰되어 다른 분류군과 명확히 구별되었다(Fig. 3J−K).
Type 3. 선모(glandular trichome)
다세포성 선모로 자루(stalk)의 유무로 두 서브타입으로 구분된다.
subtype 3-1. 짧은 자루의 두상선모(capitate glandular trichome): 20 μm 이하의 짧은 자루를 지닌 두상선모로 Chamaebatiaria속의 향축면, 배축면 모두에서 관찰되었고(Fig. 2C), S. tomentosa var. angustifolia (Fig. 3M)을 비롯한 Sorbaria속 일부 분류군에서 확인되었다. 특히, S. arborea var. subtomentosa, S. grandiflora, S. rhoifolia (Fig. 3A−B), S. sorbifolia var. stellipila (1)에서 높은 빈도로 관찰되었다.
subtype 3-2. 자루가 없는 두상선모(sub-sessile glandular trichome): 자루가 없이 잎 표면에 납작하게 위치한 두상선모로 Gillenia stipulata, Sorbaria grandiflora (Fig. 2L), S. rhoifolia (Fig. 3A−C)의 배축면에서 관찰되었고, S. arborea var. subtomentosa (Fig. 2J), S. sorbifolia var. stellipila (1)(Fig. 3G−I), S. tomentosa var. angustifolia의 향축면, 배축면 모두에서 관찰되었다. 특히, S. sorbifolia var. stellipila (1)에서 높은 빈도로 나타났다.
표피상납질
연구된 분류군 가운데 일부 분류군의 표피세포에서 표피상납질이 나타났다. Lyonothamnus속의 배축면, 향축면에서는 막성 판(membraneous platelets) 모양의 표피상납질이 확인되었고(Fig. 4A), Spiraeanthus속의 향축면(Fig. 4F), Sorbaria속의 S. arborea var. arborea (Fig. 4B), S. arborea var. subtomentosa (Fig. 4C), S. kirilowii의 향축면(Fig. 4D), S. tomentosa var. tomentosa의 배축면(Fig. 4E)에서 판(platelets) 모양이 관찰되었다.
고 찰
쉬땅나무족과 연관 분류군인, Gillenia속, Lyonothamnus속에 해당되는 20분류군에 대하여 잎 표피 미세형태학적 형질(표피세포 유형, 기공의 유형, 기공의 크기, 수층벽의 형태, 모용의 유형, 표피상납질의 유무 및 형태)을 관찰하고 기재 하였다. 본 분류군에서는 표피세포, 기공, 모용의 형태와 표피상납질의 유무 및 종류 형질에서 다양성을 관찰하였고, 형질의 유용성을 확인하였다.
연구된 분류군은 양면기공엽과 이면기공엽 두 가지 형태가 모두 관찰되었으며, 대부분 불규칙형 혹은 불규칙형과 방사형이 함께 나타났고, 드물게 다륜형이 나타났다. 일반적으로 기공은 잎의 배축면에만 나타나는 이면기공엽 형태를 보이나, 양면기공엽을 나타내는 경우도 있으며, 이 때에는 배축면의 기공 빈도가 향축면보다 높은 것으로 알려져 있다(Esau, 1977). 양면기공엽을 갖는 Adenostoma속, Chamaebatiaria속, Spiraeanthus속에서 기공의 분포가 배축면이 향축면에 비해 빈도가 다소 높은 것으로 나타나 일반적인 양면기공엽의 기공 분포를 나타내는 것으로 확인되었다. Song and Hong (2014)은 본 분류군의 잎의 해부학적 연구에서 Sorbaria속에서만 이면기공엽이 나타나는 것으로 판단하였는데, 본 연구 결과 Lyonothamnus속 역시 이면기공엽을 지니는 것으로 확인되었다. 이는 일부 단편에서 확인되는 향축면의 표피세포를 기공으로 잘못 인식한 것으로 잎 표피 형질에 대한 연구 시 주사전자현미경을 이용한 미세형태학적 연구와 광학현미경을 이용한 해부학적 연구가 동시에 수행되어야 보다 확실한 해석에 이를 수 있는 것으로 보인다.
피자식물군 내에서 기공복합체의 유형에 대한 계통진화학적 해석은 아직까지 불분명한 것으로 보인다(Cronquist, 1981). 다만, 일반적으로 불규칙형이 부세포가 없는 가장 단순하고 원시적인 유형으로 알려져 있고, 피자식물군 내에서 가장 원시적이라 생각되는 그룹인 목련목(Magnoliales)에서 주로 나타나는 평행형을 근거로 피자식물군에서 평행형이 가장 원시적인 유형으로 판단하는 경우도 있다(Baranova, 1972). Potter et al. (2007)이 제시한 장미과 계통수에서 조팝나무아과 내 가장 기저에 위치한 Lyonothamnus속에서 불규칙형이 나타나고, 쉬땅나무족 분류군보다 기저에 위치한 Gillenia속에서도 불규칙형이 나타나 기존의 견해와 일부 일치하는 것으로 나타났다. 다만, 쉬땅나무족 내 다른 분류군에서도 불규칙형이 나타나고, 기공유형의 변이가 서식환경에 의해서도 영향을 받는다는 결과가 있기 때문에(Esau, 1977), 서식지의 환경적 요인을 고려한 연구가 수행되어 기공유형의 형질이 계통진화적으로 해석 가능한 형질인지 파악할 필요가 있다. 또한, 기공의 유형 및 패턴이 식물의 진화와 서식지 환경의 이용 등의 이해를 위해 주요한 역할을 하는 바, 발생학적, 형태학적, 진화적, 생태학적, 생리학적 등 각기 다른 관점에서 해석되는 이들 형질에 대해 종합적인 고찰이 필요할 것이다(Croxdale, 2000).
공변세포의 크기와 빈도는 식물체의 유전체(genome), 생육형(growth form), 서식지 환경(environment) 등과 연관이 있는 것으로 알려져 있다. 특히, 초본은 관목과 목본 식물에 비해 유전체 크기가 크고 공변세포, 표피세포의 크기가 큰 반면에 기공의 빈도는 낮은 것으로 알려져 있다(Beaulieu et al., 2008). 또한, 건조한 환경에서는 크기가 작은 기공을 통해 수분스트레스에 따라 빠르게 반응하는 반면, 높은 빈도의 기공으로 광합성을 하는 동안 이산화탄소 확산을 최대로 하는 것으로 알려져 있다(Aasamaa et al., 2001; Hetherington and Woodward, 2003). 본 연구 분류군에서도 기공의 크기와 빈도가 음의 상관관계를 나타냄을 확인할 수 있었다. 또한, 비교적 높은 고도(1400−3800 m)의 인도, 파키스탄 등지의 서부 히말라야 일대에 분포하는 것으로 알려진 Sorbaria tomentosa var. tomentosa (Rahn, 1989)의 경우, 가장 작은 공변세포를 나타냈고, 상대적으로 그 빈도는 높게 나타났다. 반면, 가장 큰 공변세포로 나타난 Adenostoma fasciculatum var. obtusifolium을 포함한 Adenostoma속의 경우 기공의 빈도는 낮았으며, 미국의 캘리포니아주, 멕시코의 바하칼리포르니아의 산비탈에 자생하는 식물로 알려져 있다(Jepson, 1993). 추후 기후 및 서식지 생태와 기공, 표피세포의 크기와의 연관성을 비롯하여, 기후 변화에 따른 식물체의 미세형태, 패턴의 관계성 및 지표 가능성에 대한 연구가 필요할 것으로 판단된다.
본 연구 분류군에서는 크게 4종류의 모용이 확인되었고, 모용의 유형은 족 내 속간, 종간 진단형질로서 유용함이 밝혀졌다. 장미과 내 일부 분류군에서도 모용의 유용성은 이미 여러 차례 검증된 바 있다(Eriksen and Yurstev, 1999; Kolodziejek, 2008; Faghir et al., 2010; Ganeva and Uzunova, 2010; Heo et al., 2013; Faghir et al., 2014). 특히, 유선상으로 꼬인 긴 단모(subtype 1-3)는 Lyonothamnus속에서만, 농포성 선모(type 4)는 Adenostoma sparsifolium, Spiraeanthus schrenkianus에서만 나타나 종 동정을 위한 유용한 형질로 판명되었다. Rahn (1989)은 쉬땅나무속(Sorbaria)의 분류학적 재개정 연구에서 모용의 종류를 기재하고 유용성을 확인한 바 있다. 본 연구에서도 쉬땅나무 속 내 분류군에서 모용의 종류뿐 아니라 밀도의 차이 역시 유용한 형질로 판단되었다. 식물체 잎의 털의 밀도는 유전적인 영향과 환경적인 영향을 받는 것으로 알려져 있으며(Ramesar-Fortner et al., 1995; Roy et al., 1999; Sunami et al., 2013), 종간 또는 종내의 모용의 밀도는 다양한 변이로 발생할 가능성이 있다고 밝혀진 바 있다(Heo et al., 2013).
Sorbaria sorbifolia complex의 한 분류군이며 국내에만 자생하는 것으로 알려진 점쉬땅나무(Sorbaria stellipila Schneid var. glandulosa Nakai)는 Nakai가 잎 뒷면에 선점이 분포한다는 특징으로 鮮滿實用林業便覽 (Kaburaki, 1940) 에 발표하였고, 후에 Lee (1980)가 대한식물도감에 Sorbaria sorbifolia f. glandulosa로 기재하였다. 하지만, 본 학명은 비합법명으로 재고된 상태이기 때문에(Chang and Kim, 2002; Kim et al., 2005), 본 연구에서는 S. sorbifolia var. stellipila (1)의 학명을 택해 관찰하였다. Song and Hong (2014)의 해부학적 연구에서 본 분류군의 잎 배축면에 나 타나는 높은 밀도의 선점의 분포가 나머지 분류군과 확연 히 구분된다고 밝혔고, 본 연구 역시 짧은 자루의 두상선 모(subtype 3-1)와 자루가 없는 두상선모(subtype 3-2)가 S. sorbifolia var. sorbifolia의 다른 종 하 분류군들에 비해 그 빈도가 상당히 높은 것으로 확인되었다. 또한 쉬땅나무속의 수리분류학적 연구에서도 단위면적 당(0.05 mm2) 선점 의 개수가 가장 많은 것으로 나타나(Song and Hong, inprep.), 향후 형태학적, 미세형태학적 연구 결과를 종합하여 점쉬땅나무의 분류학적 계급설정 및 학명의 정당공 표가 이루어져야 할 것이다.
표피상납질은 식물군에 따라 다양한 형태로 나타나며, 분류학적으로도 매우 유용한 형질로 밝혀져 있다(Wilkinson, 1979; Barthlott et al., 1998; Ensikat et al., 2006). 또한, 장미목(Fehrenbach and Barthlott, 1988), 장미과의 일부 분류군을 포함한 연구도 꾸준히 이루어졌다(Neinhuis and Barthlott, 1997; Wissemann, 2000; Jetter and Schffer, 2001; Gostin and Adumitresei, 2010; Heo et al., 2013; Faghir et al., 2014). 본 연구에서는 Lyonothamnus속에서만 막성 판(membraneous platelets) 모양의 표피상납질이 나타나 구별 되었고, Sorbaria속 일부 분류군에서는 판(platelets) 모양이 나타나 종간 구분이 가능하였다. 표피상납질은 서식지의 환경적, 생태적 영향을 받을 뿐 아니라(Agrawal et al., 2009), 유전적인 영향 또한 받을 수 있다고 보고된 바 있다(Wissemann, 1998). 따라서, 추후 서식지 환경 특성뿐만 아니라, 염기서열에 근거한 분자계통분석 결과와도 연계시켜 해석해 볼 필요가 있다. 표피상납질에 대한 연구는 분류학적 유용성에 대한 연구와 더불어 발수성(water-repellent), 외부 오염원으로부터의 자정성(self-cleaning) 등의 기능적인 연구 또한 이루어 지고 있으며(Neinhuis and Barthlott, 1997), 식물체의 잎과 줄기를 포함한 표피상납질의 분포에 대한 연구도 이루어졌다(Tomaszewski and Zieliñski, 2014). 추후, 본 분류군 내 잎 표피 이외의 다른 영양기관과 열매에서 나타나는 표피상납질의 유무 및 종류와 더불어 기능적인 측면을 고려한 연구가 필요할 것이다.
잎의 미세형태학적 형질을 통해 최근 본 족의 분류체계를 지지하는 공유파생형질은 찾을 수 없었다. 하지만, 양면 혹은 이면 기공엽의 형태가 속 내에서 일관성 있게 나타남을 확인하였고, 기공의 크기와 빈도 및 서식지 환경과의 상관관계를 유추할 수 있었다. 또한, 모용의 다양성을 통해 본 족 내 분류학적 유용성을 파악할 수 있었다. 특히, 이전분류체계에서 본 족에 포함되었던 Lyonothamnus속에서만 유선상으로 꼬인 단모가 나타나 분자계통학적 연구를 기반으로 한 최근 족 내 분류체계인 4속과 구별됨을 확인하였다. 한편 족 내에서 관찰된 4가지 모용 중, 다양한 유형의 성상모와 선모가 특히 쉬땅나무속 내 종간 식별형질로 매우 유용함을 확인하였다. 표피상납질 역시 족 내 일부 분류군에서만 나타나 종간 식별형질로서의 유용성이 파악되었다. 본 연구를 통해 관찰된 다양한 형태의 잎 표피 미세형태학적 형질은 외부형태학적 형질과 더불어 본 족을 이해하는데 보다 유용한 정보를 제공할 것이며 향후, 근연분류군인 국수나무족(Neillieae), 조팝나무 족(Spiraeeae)를 포함한 조팝나무아과 내 분류군의 폭넓은 연구에 기여할 수 있을 것이다.
Acknowledgements
본 연구를 위해 귀중한 표본을 대여해주신 A, C, GH, K, KH, KHUS, MO, SNUA의 각 표본관의 관계자께 감사를 드립니다. 또한 논문심사 과정 중 많은 조언과 보다 정확한 논문이 되도록 교정을 해주신 익명의 두 분 심사자들께 감사드립니다. 끝으로 본 연구과정에 여러모로 도움을 아끼지 않은 경희대학교 생물학과 식물계통분류학 연구실(KHUS)의 모든 동료들에게 고마운 마음을 전합니다. 본 연구는 한국연구재단 기본연구비[세계산 쉬땅나무족(조팝나무아과: 장미과) 및 근연분류군의 계통분류학적 연구: 외부형태학, 미세형태학, 해부학 및 분자계통학, NRF-2012R1A1A2004149]의 지원으로 수행되었으며, 이에 감사드립니다.
