A new record of Tordylium maximum (Apiaceae) from Korea
Article information

Abstract
Tordylium maximum L. (Apiaceae), native to south, south-central Europe and southwest Asia and a rare alien plant in northern Europe, was newly found in Jeollabuk-do in Korea. Tordylium is clearly distinguished from other genera by having radiating petals, filiform bracts, linear bractlets, stalked mericarps with minutely vesicular dorsal face or strigose hairs, and an annual habit. Tordylium maximum is different from other species of the genus by its mericarps with smooth margins and 10–16 rays not contracted in fruit. T. maximum grows on dry and sunny grasslands. Here, we report the first occurrence of the genus Tordylium represented by T. maximum in Korea and provide a precise description, illustration, photographs of the species, and a taxonomic key to allied taxa in Korea.
INTRODUCTION
Apiaceae Lindl. (Umbelliferae Juss.) is a large family comprised of 3,000–3,800 species in ca. 355 genera (Pimenov and Leonov, 1993; Calviño et al., 2008). Many authors have proposed the classification of family Apiaceae mainly based on fruit morphology (Koch, 1824; Candolle, 1830; Bentham and Hooker, 1867; Boissier, 1872; Drude, 1898; Calestani, 1905; Koso-Poljansky, 1916; Cerceau-Larrival, 1962). Among the numerous suggestions, Drude’s classification system is broadly accepted, dividing the family into three subfamilies (subfam. Hydrocotyloideae, subfam. Saniculoideae, and subfam. Apioideae) (Downie et al., 2010; Mousavi et al., 2020). Subfamily Apioideae takes the most of the family, with more than 70 % of the species (Drude, 1898; Pimenov and Leonov, 1993; Mousavi et al., 2020). Downie et al. (2001) conducted the molecular phylogenetic analyses for the subfamily Apioideae, and they have recognized ten monophyletic tribes (Aciphylleae M. F. Watson & S. R. Downie, Bupleureae Spreng., Careae Baill., Echinophoreae Benth & Hook.f., Heteromorpheae M. F. Watson & S. R. Downie, Oenantheae Dumort., Pleurospermeae M. F. Watson & S. R. Downie, Pyramidoptereae Boiss., Smyrnieae Spreng., and Scandiceae Spreng.) and seven additional groups within the subfamily.
In Korea, 35 genera, 78 species in four tribes and four groups are found in the wild (Kim, 2019). Several authors had conducted the phylogenetic studies of the family Apiaceae in Korea, especially on the genus levels; Dystaenia Kitag. (Choi et al., 1998), Ostericum Hoffm. (Sun et al., 2000), and Sium L. (Lee et al., 2010). However, the comprehensive study of the whole family in Korea is not conducted except by Kim (2019). Meanwhile, 13 genera 14 species of Apiaceae are reported as naturalized or casual alien plants in Korea (Kang et al., 2020a). Over the past ten years, new or unrecorded species of Apiaceae from Korea have been reported occasionally. Only three new species had been reported: Angelica reflexa from Gangwon-do (Lee et al., 2013), Peucedanum chujaense from Chuja-do Island of Jeju-do (Kim et al., 2019), and Sillaphyton podagraria from Gangwon-do (Pimenov et al., 2016). Three alien plants were newly published: Bowlesia incana from Gyeongsangnam-do (Kang et al., 2020b), Chaerophyllum tainturieri from Gyeonggi-do (Hong et al., 2012), and Pimpinella saxifraga from Jeju-do (Lee et al., 2018).
Tordylium L. in tribe Tordylieae, first recorded in 1753 by Linnaeus, acomprises 20 species distributed in Europe, Asia, North Africa, South America, and Oceania (Linnaeus, 1753; Al-Eisawi and Jury, 1988). Tordylium is characterized by radiating petals, filiform bracts, linear bractlets, stalked mericarps with minutely vesicular dorsal face, and annual habit (Al-Eisawi and Jury, 1988). Within Tordylium, two subgenera are recognized based on the pubescences of the plants, shape of calyx teeth, type of pollen grains; these are subg. Tordylium and subg. Ainsworthia Drude (Al-Eisawi and Jury, 1988).
We here report the first occurrence of the genus Tordylium in tribe Tordylieae in Korea, represented by T. maximum (Figs. 1, 2). In Korea, the closest taxa to the genus Tordylium is Heracleum L. in tribe Tordylieae (Yu et al., 2011; Park et al., 2017; Lee, 2018). T. maximum is the type species of the genus. Therefore, the genus description is represented by the species description. The Korean name for the species is ‘Eo-su-ri-a-jae-bi’, followed by the nearest species in Korea, H. moellendorffii Hance.
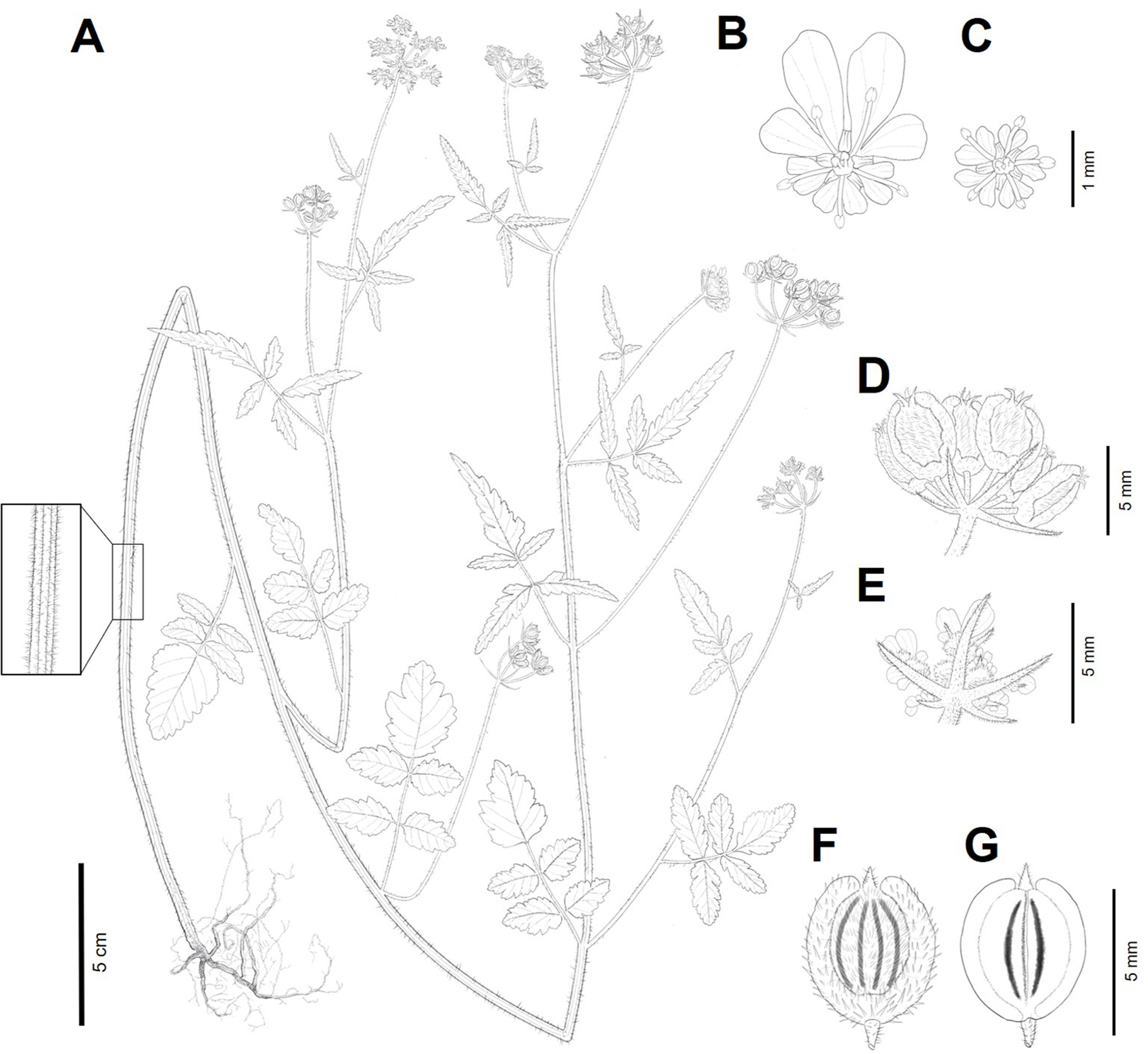
Illustrations of Tordylium maximum L. A. Habit. B. Peripheral flower. C. Central flower. D. Schizocarps. E. Bracts. F. Mericarp (adaxial view). G. Mericarp (abaxial view).

Photographs of Tordylium maximum L. A. Habit. B. Root. C. Stem. D. Cauline leaf (adaxial view). E. Cauline leaf (abaxial view). F. Compound umbel (upper view). G. Compound umbel (lower view) . H. Shizocarps. I. Mericarp (adaxial view). J. Mericarp (abaxial view).
During a field survey of plant diversity in Jeollabuk-do, an unrecorded alien plant, Tordylium maximum was confirmed in abandoned fields and roadsides near Gunsan Port, Korea. Here, we present its specific morphological description, illustrations, a taxonomic key to allied taxa as well as the photographs of its habit.
TAXONOMIC TREATMENT
Tordylium maximum L., Sp. Pl. 240, 1753 (Figs. 1, 2).— TYPE: Netherlands, without date, Clifford s.n. (Lectotype, BM! −BM000558244, designated by Al-Eisawi and Jury (1988: 390) (photo at BM through the website of https://data.nhm.ac.uk).
Korean name: Eo-su-ri-a-jae-bi (어수리아재비).
Herbs, annual or biennial, hermaphroditic, 30–80(−200) cm tall, moderately to densely pubescent with retrose setose hairs. Root a taproot, yellowish-white, branched. Stems erect, terete, branched upwardly, 0.5–0.8(−2) cm in diam., longitudinally grooved, hollow. Leaves basal and cauline, alternate, long-petiolate; petiole sheathing at base. Basal leaves caducous. Cauline leaves persistent, peiolate; petioles reduced upwardly; sheath short, triangular; blade narrowly triangular in outline, 1-pinnately compound, both surfaces densely pubescent with setose hairs, lateral leaflets sessile, terminal leaflets short petiolulate. Lower cauline leaves 6.5–13 × 5.5–7 cm, with 2 to 4 pairs of leaflets, lateral leaflets narrowly ovate to ovate, 3–3.2 × 1.2–2.2 cm, apex obtuse, base obtuse, margins irregularly crenate, terminal leaflets larger than lateral leaflets, ovate to broadly ovate, 3.5–4 × 1.6–3 cm, apex obtuse, base truncate to rounded, margins irregularly crenate. Middle cauline leaves 7.5–9.5 × 6–9 cm, with 2 pairs of leaflets, lateral leaflets lanceolate, 2.7–3.5 × 1.2–1.5 cm, apex acute, base acute, margins crenate, terminal leaflets narrowly linear-lanceolate, 3.3–5.5 × 0.7–3 cm, apex acute, base acute, margins crenate. Upper cauline leaves 4.3–8.5 × 3.7–8 cm, 3-lobed, lateral lobes narrowly oblong, 2–4 × 0.4–1 cm, apex acute, base acute, margins coarsely serrate, terminal lobe much longer than lateral lobes, linear-lanceolate, 3.6–7 × 0.5–2.3 cm, apex acute, base acute, margins coarsely serrate. Inflorescences compound umbels, terminal and lateral, flat-topped, 3–5 cm in diam.; umbellets hermaphroditic, 6–12 flowered, ca. 8 mm in diam.; peduncle up to 22 cm long; rays 6–15, ascending, 1–4 cm long, unequal in length; bracts 3–8, persistent, linear-lanceolate, entire, 0.2–1 cm long, apex acute, margins not scarious; pedicels very short; bractlets 5, narrowly linear-lanceolate, entire, 2–6 mm long, apex acute, margins not scarious. Flowers bisexual, peripheral flowers zygomorphic, central flowers actinomorphic; calyx 5-toothed, persistent, narrowly triangular, unequal in size; petals 5, white or pinkish white, obovate, ca. 0.5 × 0.5 mm, those on outer flowers of umbellets radiant, enlarged, ca. 1.2 × 1 mm, unequally 2-lobed, adaxial surface strigose; stamens 5, alternating with petals; filament filiform, ca. 1.5 mm long; anthers subglobose, 2-locular, yellow; pistil 1, 2-carpellate; ovary inferior, syncarpous, 2-locular; stylopodium conical, strigose; styles 2, 1–1.3 mm long, ascending, strigose. Fruit a dry schizocarp composed of 2 mericarps, uniform, semispherical in outline, dorsally strongly compressed; carpophore ca. 5 mm long, bifurcates to the base; mericarps suborbicular to broadly ellipsoid in outline, dorsally strongly compressed, 4–6 × 3.5– 4.5 mm, moderately to densely pubescent with strigose hairs on adaxial surface; primary ribs prominent, dorsal ribs 3, filiform, covered with strigose hairs, marginal ribs winged, wings 0.7–1 mm in width, thick, smooth or rarely corrugate; secondary ribs absent; vittae 6, 4 per vallecula, 2 on commissural face. Seed linear in cross section. Chromosome numbers 2n = 20, 22, reported by Strid and Franzen (1981).
Flowering: May to July.
Fruiting: July to August.
Distribution: Europe, Middle East, North Africa, Korea, Western Asia.
Specimens examined: KOREA. Jeollabuk-do: Gunsan-si, Osikdo-dong, 15 Jul 2020, Sang Deog Shim & Eui-ho Eom KIMJH20152 (2 sheet, KB), KIMJH20153 (KB); same locality, 5 Aug 2020, Eui-ho Eom KIMJH20154 (3 sheet, KB); same locality, 13 May 2021, Seong-Won Lee KIMJH21080 (3 sheet, KB); same locality, 17 Jul 2021, Sang Deog Shim & Eui-ho Eom KIMJH21081 (KB), KIMJH21082 (KB), KIMJH21083 (KB), KIMJH21084 (KB); same locality, 23 Jul 2021, Eui-ho Eom KIMJH21085 (KB), KIMJH21086 (KB), KIMJH21087 (KB), KIMJH21088 (2 sheet, KB); same locality, 10 Aug 2021, Kyeonghee Kim & Hwa-Jung Suh 21101 (KB), 21102 (KB), 21103 (KB), 21104 (KB), 21105 (KB).
A key to Tordylium maximum and allied taxa in Korea
1. Perennial; leaf sheath inflated; terminal leaflets 3–5-lobed; umbellets polygamous; stylopodium glabrous; vittae extending 1/3–2/3 mericarp length ······················· ··································· Heracleum moellendorffii 어수리
1. Annual or rarely biennial; leaf sheath not inflated; terminal leaflets not lobed; umbellets hermaphroditic; stylopodium strigose; vittae extending for the full mericarp length ······ ································ Tordylium maximum 어수리아재비
Notes: Tordylium maximum species was found at bare lands near roadway growing along with other herbaceous plants such as Equisetum arvense L., Chenopodium album L., Achyranthes bidentata var. tomentosa (Honda) H. Hara, Arenaria serpyllifolia L., Stellaria media (L.) Vill., Lepidium virginicum L., Amphicarpaea bracteate subsp. edgeworthii (Benth.) H. Ohashi, Medicago lupulina L., Oenothera biennis L., Oenothera laciniate Hill, Acalypha australis L., Oxalis dillenii Jacq., Metaplexis japonica (Thunb.) Makino, Solanum nigrum L., Paederia foetida L., Artemisia indica Willd., Bidens pilosa L., Erechtites hieracifolia (L.) Raf. ex DC., Erigeron annuus (L.) Pers., Hypochaeris radicata L., Senecio vulgaris L., Sonchus brachyotus DC., Sonchus oleraceus L., Bromus rigidus Roth, Digitaria ciliaris (Retz.) Koeler, and Lolium perenne L.
About 200 individuals of Tordylium maximum are discovered in open habitat, roadsides on lowlands. The population has been observed since 2018 by the authors. According to continuous observation of the population, the number of individuals is being maintained. Therefore, it is supposed the species might have adapted themselves to the local environment and the population would probably be expanded in the future. In accordance with the invasive grades categorized by Jung et al. (2016), T. maximum is considered as I (PS) grade of spread, which imply the potential invasive species.
Tordylium maximum L. belongs to subg. Tordylium sect. Tordylium and is the type species of the genus Tordylium (Al-Eisawi and Jury, 1988; Pimenov and Leonov, 1993). The sect. Tordylium contains three species that are distributed in Europe and Western Asia (Tutin, 1980; Al-Eisawi and Jury, 1988; Adams, 1999). T. maximum is most closely related to T. carmeli (Labill.) Al-Eisawi in morphology which is easily distinguished by the number of rays (10–16 vs. 3–7), lengths of the rays (less than 3 cm long vs. up to 8 cm long), shape of the rays in fruit (not contracted vs. contracted), and the sizes of the petals (3–5 mm long vs. 5–7 mm long) (Al-Eisawi and Jury, 1988).
Acknowledgements
This work was supported by a grant from the National Institute of Biological Resources (NIBR), funded by the Ministry of Environment (MOE) of the Republic of Korea (NIBR202102103, NIBR202203102).
Notes
CONFLICTS OF INTEREST
The authors declare that there are no conflicts of interest.