Perennial herbs. Stem pubescent, rarely glandular. Leaves alternate, sessile, tomentose to villose or cobwebby hair; margin entire. Capitula heterogamous, rarely dioecious, disciform, in dense or loose terminal corymbs. Bracteal leaves similar with upper cauline leaves, white to grayish or yellowish tomentose. Phyllaries usually 3-seriate, papery, monomorphic, brownish, transparent. Marginal florets yellow, pistillate, filiform, outnumbering the disk florets. Disk florets functionally male; corolla yellow, 5-lobes erect, sparsely glandular in apex of the lobes. Anthers ecalcarate, with short tails. Pollen gnaphalioid. Style bifid; style-branches truncate with densely hirsute stigmatic surface separated. Achenes glabrous to papillose or pubescent. Pappus basally connate to ring, barbellate, capillary bristles.
A key to the varieties of L. coreanum
1. Plant height 15.5–31.5 cm; cauline leaves of middle part 30.6–71.5 mm long, lanceolate, narrowly oblong, oblong; densely or loosely corymbous heads; bracteal leaves oblong-lanceolate, ovate ························· var. coreanum
1. Plant height 8.5–11.3 cm; cauline leaves of middle part ca. 2 cm long, elliptical, ovate, oblanceolate; densely corymbous heads; bracteal leaves elliptical, oval, oblaceolate ·········································· var. hallaisanense
Var. coreanum
Korean name: 솜다리 Som-da-ri
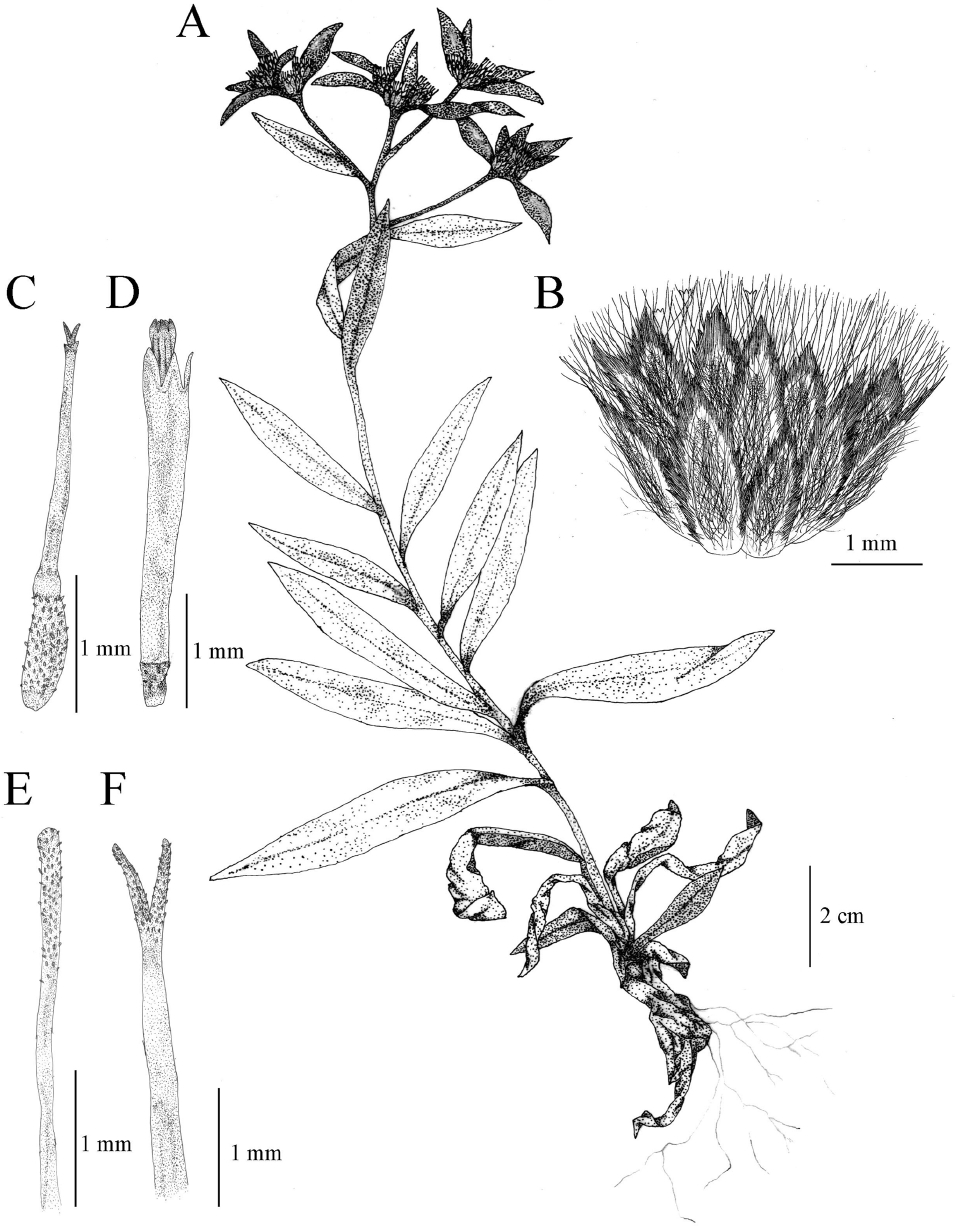
Perennial herbs. Rhizome prostrate, branching, with flowering and sterile stems. Stems erect, 15.5–31.5 cm tall, internodes 9.6–25.0 mm, lower half somewhat becoming woody, pubescent. Leaves alternate, 7–21, 30.6–71.5 mm long, 6.1–13.7 mm wide, upper surface of leaves green and thinly tomentose, lower surface of leaves whitish tomentose, lanceolate, narrowly oblong, oblong, margin entire, apex acute to acuminate, base abruptly narrowed, sessile, basal leaves deciduous at flowering periods. Inflorescence solitary or branched on flowering stems. Heads 7–19, arranged in compact or sparse terminal corymbs. Bracteal leaves 11.3–18.9 mm length, 3.3–6.6 mm wide, usually smaller than cauline leaves, oblong-lanceolate, ovate, both surfaces densely pubescent. Involucre subglobose, 4–5 mm high, 3.0–4.8 mm across; phyllaries 3-seriate, acute, margin brownish scarious, densely hairly dorsally. Marginal florets female, capillary, 5-lobed with ciliolate; stigma bifid, densely hirsute; ovary densely papillose. Disk florets bisexual, corolla subtubular, 5-lobed, yellowish, sterile; stigma densely hirsute; ovary thinly hirsute apex; anther basal sagittae. Achenes 1.2–1.6 mm long, papillose. Pappus of capillary bristle, 2.6–3.0 mm length. Flowering July–August. Fruit August–September (
Fig. 2).
Distribution: Endemic to Korea. Gangwon-do, Gyeonggi-do, Chungcheongbuk-do, Gyeongsangbuk-do. Growing on alpine meadow or forest on northward from central Korea.
Taxonomic note: The taxonomic entity of
Leontopodium coreanum Nakai has long been confused with its congener L. japonicum, which is widely distributed in East Asia. Plants of L. coreanum were first recognized by Nakai (1917) as endemic to Mt. Keumgang of Gangwon-do. Subsequently,
Nakai (1920) described its morphology as very similar to Japanese
L. japonicum, being distinguished by its leaf shape, minute amount of hairs on the ovaries of disk florets, and a liberal base for the pappus, when compared with plants of Japanese
L. japonicum. Nevertheless, those two taxa have not been previously distinguished definitively due to subtle morphological differences, and the taxonomic entity of plants in South Korea also remains unclear – whether
L. coreanum or
L. japonicum. Accordingly, those two taxa have been recorded together in most regional floras of Korea (e.g.,
Lee, 1980;
Lee, 1996;
Kim, 2007), whereas a previous monograph by
Handel-Mazzetti (1928), as well as floras for China (
Chen et al., 2011) and Japan (
Koyama, 1995) restricted the distribution of
L. japonicum to those two countries.
In this study, our morphological comparisons suggested that the plants of South Korea are closely related to
L. coreanum, and should be treated as an independent species rather than conspecific with
L. japonicum of China and Japan, based on its diagnostic characters (
Table 2). Although the character for the liberal base of the pappus, generally observed in other genera (e.g.,
Anaphalis DC. and
Pseudognaphalium Kirp.), is not adequate for species of
Leontopodium, the other morphological characters for populations in South Korea are similar to those of the type locality, Mt. Keumgang. In particular, the character for hairs on the ovaries of disk florets distinguishes plants of South Korea as well as
L. coreanum from
L. japonicum (
Table 2). In contrast to a previous proposal by
Lee and Park (2008), which treated
L. coreanum as a synonym of
L. japonicum based on the glabrous achenes in both taxa, we determined that
L. coreanum, which has hairs on the ovaries of the disk florets during fruiting, is distinct from the glabrous samples of
L. japonicum that we examined here. Furthermore, the taxonomic boundary of
L. coreanum is clearly supported by our previous cytological study (Korea: 2n = 28; Japan: 2n = 26;
Lee et al., 2010) and by molecular data for genetic divergence (China and Japan: 0.471, Korea and China: 0.742, Korea and Japan: 0.846, unpubl. data), thereby implying differentiation of the Korean populations. Based on this evidence, we suggest that
L. coreanum, closely related to
L. japonicum, is in the process of differentiation (e.g,, allopatric speciation due to oceanic barriers), and must be accepted as an independent species in accordance with a treatment by
Nakai (1917). However, when considering the regional morphological variations (China: var.
saxatile, var.
microcephalum; Japan: var.
perniveum, var.
shiroumense, var.
spathulatum) as well as the wide distributional ranges of
L. japonicum, further details, e.g., molecular and micro-morphological data for their phylogenetic relationships, are needed. Although
Handel-Mazzetti (1928) treated
L. coreanum as a synonym of
L. discolor, without discussion, several characteristics of the former, including its oblanceolate cauline leaves and glabrous achenes, provide sufficient distinction from
L. discolor (lineari-ovate cauline leaves and pubescent achenes), as found in our morphological examination. For now, we are making provisional decisions about those South Korean plants as belonging to
L. coreanum, due to their similarities in diagnostic characters. In the protologue,
Nakai (1917) indicated three original materials. We identified two of the original materials at TI, and selected one (
T. Nakai 5915) having complete information in accordance with the protologue as lectotype (ICN Art. 9.2, 9.5, 9.12,
McNeill et al., 2012).
Var. hallaisanense (Hand.-Mazz.) D.H. Lee & B.H. Choi, comb. stat. nov.
Leontopodium hallaisanense Hand.-Mazz. Beih. Bot. Centralbl. 44: 72. 1928. Type. Korea, Jeju Isl. Aug. 1907,
U. Faurie 1964 (Lectotype, designated here, E, see
Fig. 3). Remaining syntype. Korea, Jeju Isl. Aug. 1907,
U. Faurie 1967 (LE, seen as photo!; G, seen as photo!).
Korean name: 한라솜다리 Hal-la-som-da-ri
Stems erect, 8.5–11.3 cm tall, pubescent, in lower part often somewhat ligneous. Leaves alternate, 9–11, 19.2–20.8 mm long, 5.0–7.2 mm wide, elliptical to ovate, oblanceolate. Inflorescence usually solitary on flowering stems. Heads 7–8, arranged in compact terminal corymbs. Bracteal leaves 13.5–16.3 mm length, 5 mm wide, elliptical, oval, oblanceolate. Involucre subglobose, 5.0–5.5 mm high, 4.5–5.5 mm across (
Fig. 4).
Distribution: Endemic to Korea. Growing on summit of Mt. Halla on Jeju Isl.
Taxonomic note: Var.
hallaisanense is currently restricted to the upper part of Mt. Halla, where only a few individuals (n<30) occur. These plants were originally described by
Handel-Mazzetti (1928) as being an independent species that differed from
L. japonicum based on plant height and shape of the bracts (
Fig. 3). However, those characters are commonly found for
Leontopodium (
Ling, 1965), and some authors have questioned the validity of having a separate species (
Lee, 1996;
Lee et al., 2010). Indeed, such diagnostic characteristics have also been reported for several specimens from inland populations of
L. coreanum, even though such types are rarely identified. Thus, it might not be sufficient to recognize it as an independent species. Although the number of examined specimens for
L. hallaisanense has been limited, the results of our molecular analysis (Lee et al., unpubl. data) demonstrated that
L. hallaisanense was nested within
L. coreanum. Therefore, we suggest that var.
hallaisanense be considered an ‘insular form’ of
L. coreanum, differentiated by diagnostic characteristics as key, and should be treated as a variety of that species. Furthermore, previous reports of the distribution of
Leontopodium species at Mt. Halla [
L. japonicum: upper area of Mt. Halla (
Nakai, 1913);
L. coreanum: 1800–2100 m (
Nakai, 1920)] may have led to a misidentification of var
. hallaisanense. Therefore, further ecological and molecular analyses with more individuals are necessary if we are to develop a conservation strategy for the few plants (n<30) that remain at the peak of Mt. Halla.
2. Leontopodium leiolepis Nakai in Matsumura, Icon. Pl. Koisikav. 4: 75. 1920. Type. Korea, Hamgyeongnam-do, Waigalbong, 2,200 m, Jul. 11, 1914,
T. Nakai 1542 (Holotype, TI!, see
Fig. 5).
Leontopodium seorakensis Lim, Hyun, Kim & Shin, Korean J. Pl. Taxon. 42: 157. 2012. Syn. nov. Type. Korea, Kangwon-do, Inje-gun, near the peak of Mt. Ahn (1,396 m) area in Mt. Seolak National Park, 1,360 m elev., N 38°08′10.2″, E 128°20′4.2″, 30 Jun. 2007, Hyun 20070004 (Holotype, KB; Isotype, SNU).
Korean name: 산솜다리 San-som-da-ri
Perennial herbs. Rhizome prostrate, branching, with flowering and sterile stems. Stems erect, 6.0–11.5 cm tall, woolly hair and glandular. Leaves alternate, 5–9, 16.0–22.8 mm long, 3.0–5.5 mm wide, upper surface of leaves thinly tomentose, lower surface of leaves whitish arachnoid and glandular, both sides becoming sometimes calvescent, spatulate, linear or oblanceolate, margin entire, apex mucronate, base attenuate, sheathing stems, radical and basal cauline leaves present at flowering periods. Inflorescence solitary on flowering stems. Heads 5–9, densely arranged. Bracteal leaves 11.3–17.3 mm length, 2.7–4.3 mm wide, more or less smaller than cauline leaves, linear or oblanceolate, upper surface densely floccose, lower surface thinly arachnoid. Involucre subglobose, 5.0–5.5 mm high, 3.5–4.5 mm across; phyllaries 3-seriate, 3.0–4.3 mm long, 1.1–1.4 mm wide, acute, margin brown to reddish brown scarious, glabrous, sparsely glandular. Marginal florets female, corolla capillary, 4-lobed with sparsely ciliolate; style cylindrical; stigma bifid, densely hirsute; ovary densely papillose. Disk florets bisexual, corolla subtubular, 5-lobed, yellowish, sterile; style cylindrical; stigma clavellate, densely hirsute; ovary pubescent; stamens 5; anther basal sagittate. Achenes 0.9–1.7 mm long, papillose. Pappus of capillary bristles, 2.4–3.1 mm length. Flowering May–June. Fruit July–August (
Fig. 6).
Distribution: Endemic to Korea. Hamgyeongbuk-do, Hamgyeongnam-do, Pyeonganbuk-do, Gangwon-do. Rock crevice on high mountains of northern and central Korea.
Taxonomic note:
L. leiolepis was identified by
Nakai (1920) from samples collected at Waigalbon (2,200 m) in North Korea, and was considered an endemic species distributed northward from Mt. Seolak in South Korea. Recently, however,
Lim et al. (2012) have described plants from Mt. Seolak as a new species,
L. seorakensis Lim, Hyun, Kim & Shin, separate from
L. leiolepis. In their protologue, they suggest that
L. seorakensis is distinguished from the latter by morphological characters such as densely arachnoid hairs on the lower surface of bracteal leaves as well as sparsely glandular outer phyllaries that are of similar size and shape as the former. However, those diagnostic characteristics were already described and illustrated in the declaration for
L. leiolepis by
Nakai (1920) as “supra floccose infra aranea” and “deorso paruissime glandulosociliata”. Indeed, specimens for
L. leiolepis, including type materials, have almost glabrous and sparsely glandular hairs on the outer phyllaries that are easily distinguished from other
Leontopodium species. Our morphological comparison between specimens from the type locality of
L. seorakensis [locotype: Mt. Ahn, Jun. 12, 2009,
D.H. Lee & W.B. Cho 96015, 96018, 96019 (IUI!);
D.H. Lee & W.B. Cho 96017,
96020 (KH!)] and those of
L. leiolepis, including the type specimen, also indicate no clear morphological discontinuities between the two species, but further support the merging of them (
Table 3). Although several specimens collected in North Korea actually exhibited somewhat glabrous bracteal leaves, such variations as density of hairs on those leaves are relatively easily recognized within the genus as intraspecific variations (
Ling, 1965). Moreover, the plant materials in both the type specimen of
L. leiolepis (
Fig. 5) and the type locality of
L. seorakensis have almost glabrous and arachnoid bracteal leaves. In fact, the glabrous bracts are mostly observed in plants during fruiting periods, and we assume that this variation in density might be influenced by the time when they were collected (i.e., flowering: arachonid; fruiting: thinly arachnoid to almost glabrous). Another morphological character for
L. seorakensis is also ambiguous.
Lim et al. (2012) have stated that the length and shape of the outer and inner phyllaries are similar in
L. seorakensis, but no exact quantitative values exist. In contrast, our examinations indicated that this species also has slightly narrow and longer inner phyllaries when compared with the outer ones, and the ratio of lengths between inner and outer phyllaries for
L. seorakensis (1.06–1.28 mm) does not differ much from that for
L. leiolepis (1.14–1.36 mm;
Table 3). Hence, we assume that the above diagnostic characters for
L. seorakensis are not sufficient to describe it as a new species that is separate from
L. leiolepis, despite their subtle differences. Rather, we might infer that those differences are just included in the intraspecific variations of
L. leiolepis (
Fig. 5) and, thus, we deduce here that
L. seorakensis is a synonym of
L. leiolepis.
Meanwhile,
L. leiolepis has been classified within the subsection
Kurilensia Hand.-Mazz., having glandular hairs on the stems that are similar to those of
L. kurilense Takeda, which is endemic to Japan (
Handel-Mazzetti, 1928). Furthermore, several varieties (i.e., var.
curvicollum and var.
crinulosum) have been proposed for populations in North Korea (
Im, 1999). However, its distribution and taxonomic relationships had not previously been evaluated. In addition, the isolated habitats of Mt. Seolak represent the South limit of its distribution range. Therefore, further molecular and population analyses are necessary if we are to gain broad knowledge about the phylogenetic relationships and conservation of this species.
3. Leontopodium leontopodioides (Willdenow) Beauverd in Bull. Soc. Bot. Geneve, Ser. 2, 1: 371. 1909. Type. Russia, Siberia, s.d.
Willdenow s.n. (Holotype, BM, seen as photo!,
Fig. 7)
Filago leontopodioides Willdenow, Phytogr. 12. 1794. Gnaphalium leontopodioides (Willdenow) Willdenow, Sp. Pl. 3: 1893. 1804. Antennaria leontopodioides (Willdenow) Nakai in Bull. Nat. Sci. Mus. Tokyo 31: 112. 1952.
Korean name: 들떡쑥 Deul-tteok-ssuk
Perennial herbs. Rhizome short, lignose, multicipital, with flowering and sterile stems, not radical leaves. Stems slender or erect, somewhat flexuous, 23–42 cm tall, covered with woolly hair, then becoming peeled. Leaves sparsely alternate, 12–22, similar in size and shape, 22.9–35.6 mm long, 3.6–6.3 mm wide, linear to lanceolate, apex rounded to mucronate, margin entire, base gradually narrowed, half-sheathing stem, upper surface greenish and thinly pubescent, lower surface densely pubescent. Inflorescence depauperate, top or axila on flowering stems. Heads 1–4, dioecious. Bracteal leaves 1–4 per heads, smaller than cauline leaves, 12.3–21.4 mm long, 3.2–6.4 mm wide, linear to lanceolate, inner phyllaries more narrowed, colorless or brownish scarious, densely hairly dorsally. Achenes 0.9–1.0 mm length. Pappus uniseriate, 3.6–4.6 mm long, grayish white. Flowering May–June. Fruit July–August (
Fig. 8).
Distribution: China, Korea, Mongolia, Russia.
Korea: Hamgyeongnam-do, Chungcheongbuk-do, Gyeongsangbuk-do, Gyeongsangnam-do. Sunny grassland on central Korea.
Taxonomic note: This species was first recorded as
Filago leontopodioides Willdenow from Siberia before being transferred to the genus
Leontopodium, i.e.,
L. leontopodioides (Willdenow) Beauverd. This treatment is now broadly accepted for plants in their distribution range of China, Mongolia, Russia, and Korea (
Grubov, 1990;
Kim, 2007;
Chen et al., 2011). A Korean distribution was first recorded in Seoul by
Palibin (1898) and assigned the name
Gnaphalium leontopodioides. It has been also collected from Chinampo in the northern region of Korea (
Beauverd, 1912;
Handel-Mazzetti, 1928). Compared with other species in that genus,
L. leontopodioides grows at relatively lower elevations (300–700 m) and has a wider distribution range. This taxon, being dioecious, is well-distinguished by inflorescences positioned at the top or axillary to the flowering stems. It lacks a pedicel and pappus and its flower heads are 1.5 times longer than those of other
Leontopodium species.