The distribution of Hong-do-go-deul-ppae-gi was first recognized in Korea by identifying the specimen (Ishidoya, T. & T. H. Chung 3741 TI) collected by T. Ishidoya on Hongdo Island as Paraixeris denticulato-platyphylla (Makino) Nakai, a hybrid of Paraixeris denticulate Nakai [= Crepidiastrum denticulatum (Houtt.) J. H. Pak & Kawano], and C. lanceolatum var. latifolium Nakai [= C. platyphyllum (Franch. & Sav.) Kitam.]. Later, it was treated as ×Crepidiastrixeris denticulato-platyphylla (Makino) Kitam. by Kitamura (1937). It seems to have been named Hong-do-go-deul-ppae-gi because it was first collected on Hongdo Island (Lee, 1969; Lee, 1996). However, the above scientific name was illegitimate (ICN H6.2. and H.9.1.). Hence, it was corrected to Crepidiastrum ×nakaii H. Ohashi & K. Ohashi, as the two parent species were treated as Crepidiastrum by Pak and Kawano (1992) (Ohashi and Ohashi, 2007). Saito et al. (2006) reported that C. ×nakaii occurs in the area where the parents occur sympatrically, and C. ×nakaii shows repeated frequent hybridization between the parent taxa as a result of an incomplete breeding barrier between the two taxa. However, there is disagreement as to whether or not C. platyphyllum is distributed in Korea (Ohwi, 1965; Koyama, 1995; Kadota et al., 2017). Furthermore, Lee (1996) described Hong-do-go-deul-ppaegi as a hybrid of C. denticulatum and C. lanceolatum. Therefore, controversy exists regarding the identity of this plant.
Natural hybridization is a frequent evolutionary phenomenon in flowering plants (Grant, 1981), playing an important role in the formation of new species (Arnold, 1997; Rieseberg, 1997; Seehausen, 2004). Under the limited migration from the continent, species richness of insular flora can be increased via hybridization which may lead to the generation of new species with novel combinations of potentially advantageous parental traits in the new environment (Francisco-Ortega et al., 1996; Brochmann et al., 2000; Jorgensen and Olesen, 2001; Fuertes-Aguilar et al., 2002; Howarth and Baum, 2005). Because of the importance of plant hybridization in evolution, it is of great importance to accurately identify hybrid individuals.
To determine the taxonomic identity of Hong-do-go-deul-ppae-gi, this study examined its morphological characteristics and distribution, and analyzed nuclear ribosomal DNA internal transcribed spacer (ITS) sequences with its related species in the genus Crepidiastrum.
Materials and Methods
Morphological observation and survey of geographical distribution
To compare the morphological characters of Hong-do-go-deul-ppae-gi and its closely related taxa, we collected specimens from Hongdo, Jindo, and Geomundo Islands in Jeollanam-do Province, South Korea, and deposited them in the Herbarium of Inha University (IUI). In addition, we examined additional specimens throughout the Korea National Biospecies Information System (http://www.nature.go.kr). For identification, reference was made to Kitamura (1937, 1955), Pak and Kawano (1992), and Ohashi and Ohashi (2007). Distribution data were collected from the Global Biodiversity Information Facility (GBIF) (http://www.gbif.org), the Korea National Biospecies Information System (http://www.nature.go.kr), Science Museum Net (http://science-net.kahaku.go.jp), and the Chinese Virtual Herbarium (http://www.cvh.ac.cn).
DNA extraction and PCR amplification
Total genomic DNA was extracted from dried leaf materials with silica gel (deposited in the herbarium of Inha University, IUI) using the DNeasy Plant Mini Kit (QIAGEN, Hilden, Germany) according to the manufacturer’s instructions. The extracted DNA was electrophoresed on a 1% agarose gel to confirm the presence or absence of DNA, and the concentration and quality of DNA were confirmed using a NanoDrop ND-1000 (NanoDrop Technologies, Wilmington, DE, USA). We used PCR amplification of the ITS of the nuclear ribosomal DNA. Primer and PCR conditions were as described in Saito et al. (2006). The PCR products were visualized on 1% agarose gels and sequenced using an ABI 3730xl DNA analyzer (Applied Biosystems, Foster City, CA, USA). The determined sequences were deposited in the Genbank (accession nos. MW938634–MW938646). Additionally, the nuclear ribosomal ITS sequences of Japanese and Chinese individuals deposited in GenBank were used for the ITS analysis (Table 1).
Phylogenetic analysis
The analyzed nucleotide sequences were determined after checking the chromatogram using Geneious R 7.1.9 (Biomatters Ltd., Auckland, New Zealand). All sequences were aligned using the MUSCLE software (Edgar, 2004). Maximum likelihood trees based on ITS data were constructed using the W-IQ-TREE Server (http://iqtree.cibiv.univie.ac.at/) (Trifinopoulos et al., 2016), based on user-friendly web servers for IQ-TREE 1.5 (Nguyen et al., 2015). Each aligned sequence dataset was tested to determine the best-fit model by using W-IQ-TREE with the Akaike criterion, and new model selection procedures. TIM3e was confirmed as the best-fit model for the ITS. We evaluated the node supported by 1,000 ultrafast bootstrap replicates (UFBS) (Minh et al., 2013). As an outgroup, Youngia japonica (L.) DC., which is most closely related to the genus Crepidiastrum, was used (Kilian et al., 2009).
Results and Discussion
Observation of morphological traits
According to Pak and Kawano (1992), C. denticulatum is a herb, lacking radical leaves during the flowering season, membranous cauline leaves with acute serrations, 13–15 florets, and short and shout beaked achenes. In contrast, C. lanceolatum and C. platyphyllum are subshrubs, persisting radical leaves, and thicker cauline leaves with entire and beakless achenes. In addition, C. lanceolatum has 8 inner involucre bracts, 8–12 florets, and C. platyphyllum has 5 inner involucre bracts, and 5 florets. As a result of observations, Hongdo Island individuals showed mixed characteristics of C. denticulatum and C. lanceolatum (Fig. 1, Table 2). Hongdo Island individuals were similar to C. lanceolatum in a shrub with caudex stout, 8 inner involucre bracts, florets less 10–12 (13–15 in C. denticulatum), but differed from C. lanceolatum by having radical leaves withered during the flowering season, membranous cauline leaves with acute serrations, and short and shout beaked achenes as C. denticulatum.
Survey of geographical distribution
To confirm the distribution of Hong-do-go-deul-ppae-gi and its related taxa, Hong-do-go-deul-ppae-gi was collected from Hongdo and Geomundo Islands in Korea. Crepidiastrum lanceolatum occurs in rocky areas along the seashore of Jeju and the southern coast of Korea, while C. denticulatum occurs on sunny slopes and roadsides in the mountains of all Korean provinces. According to recent literature, C. platyphyllum is known as a Japanese endemic plant distributed in Chiba Prefecture, Kanagawa Prefecture, and Shizuoka Prefecture in Japan (Pak and Kawano, 1992; Koyama, 1995; Kadota et al., 2017). Therefore, it is highly likely that a hybrid of C. lanceolatum and C. denticulatum, rather than C. platyphyllum and C. denticulatum, would occur on Hongdo Island (Fig. 2). In addition, the T. Ishidoya specimen (Ishidoya, T. & T. H. Chung 3741 TI) was re-identified as a hybrid of ×Crepidiastrixeris denticulato-lanceolata Kitam. (nom. illeg.) [= C. ×muratagenii H. Ohashi & K. Ohashi] by Kitamura (1955).
Phylogenetic analysis of ITS sequences
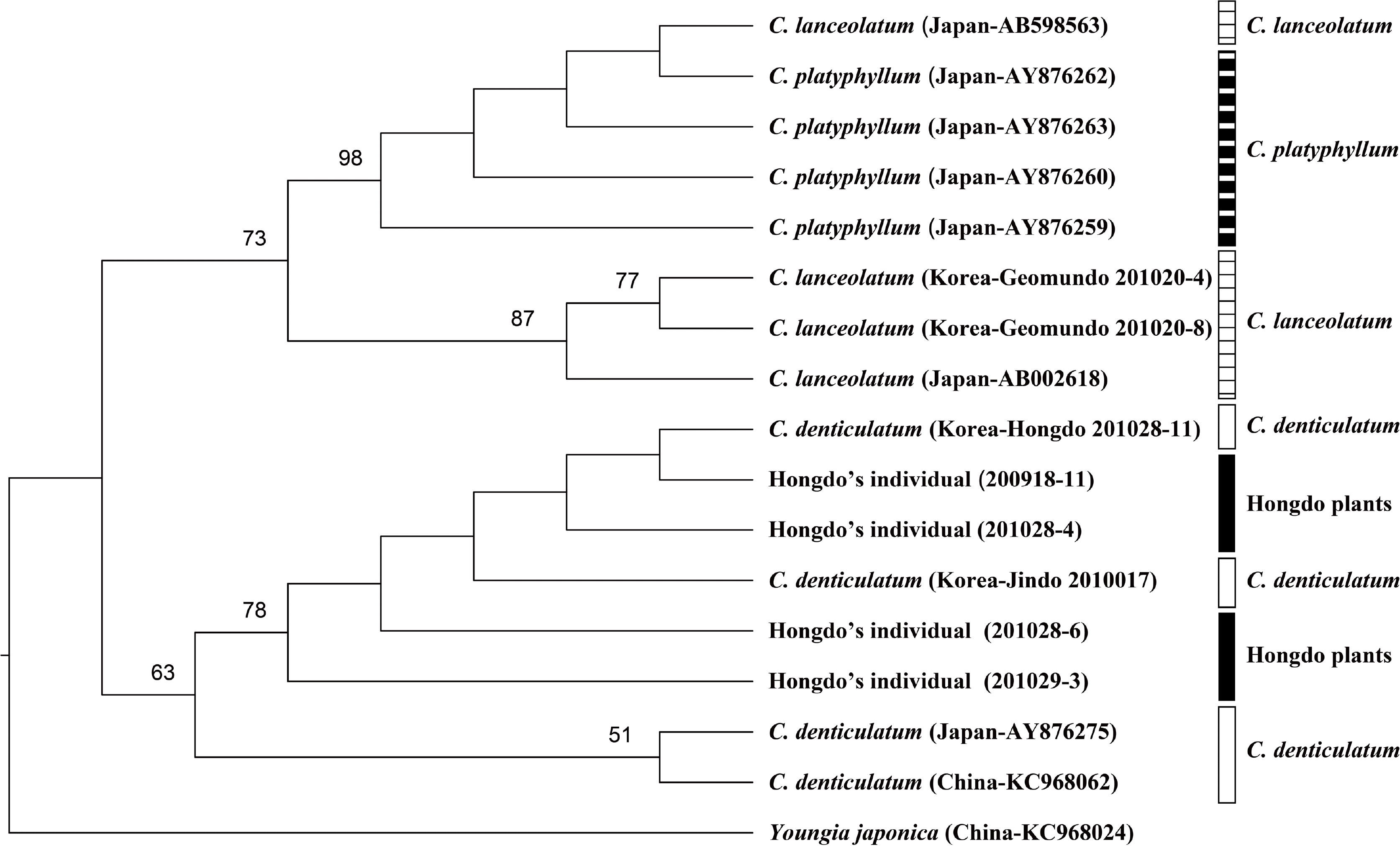
In the ITS phylogenetic tree, the 4 taxa of the genus Crepidiastrum were divided into two clades; C. lanceolatum and C. platyphyllum, C. denticulatum, and Hongdo Island individuals, but each taxon did not form a monophyletic group. Significant differences did not exist between the nucleotide sequences of the Hongdo Island individuals and the Korean C. denticulatum (Fig. 3). For that reason, Saito et al. (2006) reported that C. ×nakaii in F1 status showed an additivity of parent sequences at the nrDNA locus, but in F2 status showed either the ITS sequences of one parent only, or one predominant ITS type, despite the intermediate morphology between C. platyphyllum and C. denticulatum. These results can be explained through a backcross to C. denticulatum or segregating of F2. Further studies are required to understand the taxonomic identity of this species.
Origin and scientific name of the plant
Based on our investigations on morphological traits and geographical distribution, the origin of “Hong-do-go-deul-ppae- gi” recorded in Hongdo Island should be regarded as a hybrid between C. denticulatum and C. lanceolatum, and not a hybrid between C. denticulatum and C. platyphyllum. Therefore, that the scientific name of the plant should be assigned as C. ×muratagenii, a hybrid between C. denticulatum and C. lanceolatum.