RESULTS AND DISCUSSION
Chaenorhinum (DC.) Rchb., Consp. Regn. Veg. 123, 1828
Korean name: Ja-gal-pul-sok.
Herbs annual or perennial. Stems erect, simple or branched, glandular hairs, sometimes glabrous. Leaves opposite at lower, alternate at upper, sessile, linear or oblanceolate, margin entire.
Inflorescences axillary or solitary. Flowers bisexual, zygomorphic; calyx 2–5-lobed; corolla bilabiate, spurred; upper lip 2-lobed, lower lips 3-lobed; stamen 4, didynamous; pistil 1. Capsules ovoid to subglobose, poricidal dehiscence. Seeds 40–60, ovoid to truncate-conical, brown to blackish brown, ribbed longitudinally.
Distribution: Europe, North America, North Africa, Southwest Asia, East Asia (Russia, China, Japan, Republic of Korea).
Chaenorhinum minus (L.) Lange, Prodr. Fl. Hispan. 2: 577, 1870 (
Figs. 2–4).
Antirrhinum minus L. Sp. Pl. 2: 617, 1753.
Korean name: Ja-gal-pul.
English name: Dwarf snapdragon.
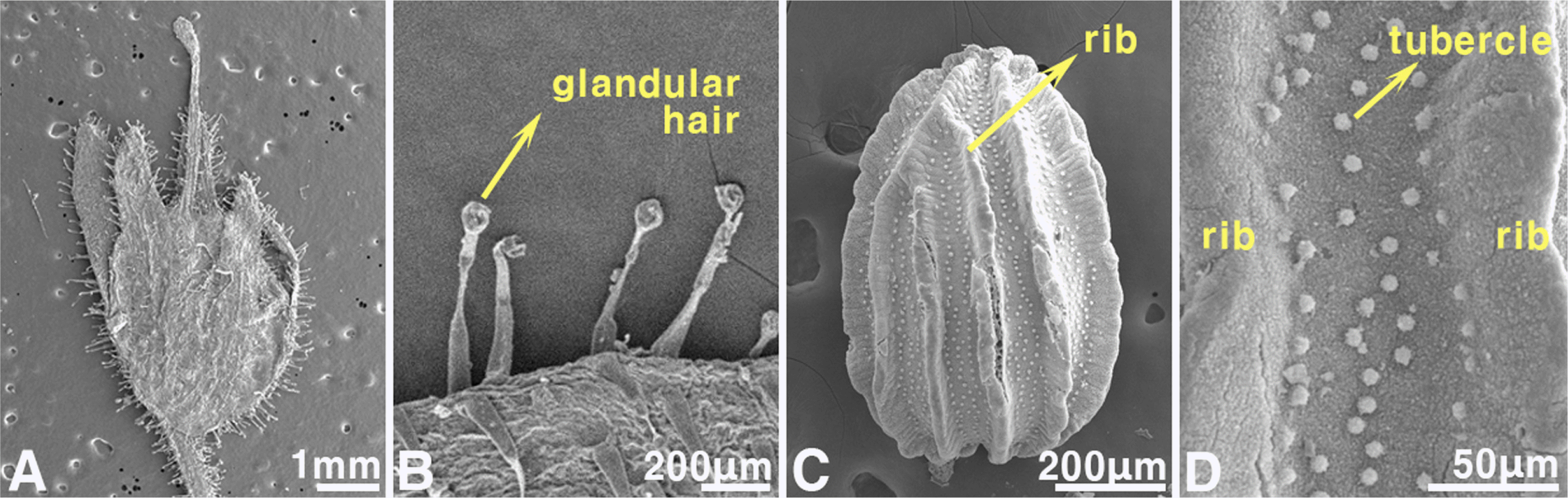
Herbs annual, 17.9–24.5 cm tall. Stems monomorphic, fertile, erect, simple or branched, yellowish green or purplish brown at base, glandular hair. Leaves opposite at lower, alternate at upper, sessile, 1–2.3 cm × 1.3–4 mm, linear, oblanceolate, spatulate, apex acute or obtuse, margin entire, base attenuate, green, glandular hair, slightly recurved. Inflorescence axillary; pedicel 9.6–14.1 mm long, yellowish green, glandular hair. Flowers bisexual, zygomorphic; calyx 5-lobed, 1.7–3.3 × 0.3–0.7 mm, linear or oblong, apex acute or obtuse, margin entire, yellowish green, glandular hair; corolla bilabiate, spurred, 5.6–7.9 mm long including spur; tube tubular, 2–4.5 mm diam., whitish purple to purple, glabrous on inside, hair on outside; upper lip 2-lobed, 1.5– 2.6 mm long, pale purple to purple at adaxial surface, glabrous on inside, hair on outside; lower lip 3-lobed, 1–2 mm long, white, slightly hairs on palate, glabrous on inside, hair on outside; spur 1.2–1.6 mm long, conical, pale purple to purple; stamens 4, didynamous; filament linear, longer filament 2.5– 4.3 mm long, shorter filament 1.3–2.7 mm long; anther 0.6– 0.8 mm long, ellipsoid, purple, dorsifixed; pistil l, ovary 1.4– 1.6 mm long, ellipsoid, purple, glandular hair; style 2.5– 2.7 mm long, filiform, glandular hair at base, persistent. Capsules 4.8–5.5 × 3.3–4.4 mm, ovoid, yellowish green or reddish brown, glandular hair, poricidal dehiscence. Seeds 0.7–1 × 0.5–0.7 mm, widely ovoid or oblong, brown to blackish brown, ribbed longitudinally, tuberculate between ribs.
Flowering and fruiting: May to October.
Origin: Europe, North Africa.
Distribution: Europe (
Fernandes, 1971,
1972), North Africa (
Davis, 1978a,
1978b;
Benedí and Güemes, 2009), North America (
Arnold 1991;
Güemes et al., 2016), East Asia (Russia, China, Japan, Republic of Korea) (
Craig and Richard, 2019;
Kim et al., 2019;
POWO, 2024).
Distribution in the Republic of Korea: Chungcheongnam-do (Dangjin-si).
Specimens examined: Chungcheongnam-do, Dangjin-si, Songsan-myeon, Gagok-ri, 28 Jun 2021, J.S. Kim KJS0844 (KH); same locality, 10 Oct 2021, J. S. Kim KJS1004, KJS1005 (KH); same locality, 16 Oct 2021, KJS1327 (KH); same locality, 16 Jun 2023, E. S. Kang and Y. K. Joung Gagokri-230616-009, Gagokri-230616-010, Gagokri-230616-011 (KH).
Taxonomic note: The genus
Chaenorhinum, to which
C. minus belongs, has a morphology similar to that of
Nuttallanthus among related genera in the Republic of Korea. The common features of the two genera include linear or oblong leaves, an axillary five-lobed calyx, pale purple corolla, and white lower lip palates (
Ji et al., 2012;
Conn and Murray, 2015). However, this genus differs from
Nuttallanthus as it has glandular hairs on the plant (
Figs. 3D,
4A, B); monomorphic, fertile, and erect stems; long inflorescences of 9.6–14.1 mm; a short corolla of 5.6–7.9 mm; a white base of the corolla lower lips; and widely ovoid or oblong seeds, making them easily distinguishable (
Fig. 3C–G, K).
Chaenorhinum minus is known to thrive in sunny, dry soils and to be primarily found on rocks, railroads, cultivated fields, and sandy and gravelly areas (
Fernandes, 1972;
Beckett, 2000;
Benedí and Güemes, 2009;
Starchenko and Darman, 2017). The
Chaenorhinum minus discovered in Dangjin-si, was also found growing in a gravel-covered vacant lot near a port (
Figs. 1,
3A). Referring to such growth characteristics, the assigned name of this species in Korea is ‘Ja-gal-pul.’
In 2021, approximately 40 individuals of C. minus were discovered for the first time in Dangjin-si. However, by 2023, the number had increased to approximately 100 individuals, suggesting gradual growth of the population of this species. Currently, C. minus occupied an area of around 12,000 m2 at the habitat, and in the area, there was an abundance of alien plants or cultivated plants compared to native plants. Native plants consisted of a total of nine taxa, Albizia julibrissin Durazz., Artemisia indica Willd., Aster hispidus Thunb., Bromus japonicus Thunb., Calamagrostis epigejos (L.) Roth, Lactuca indica L., Miscanthus sinensis Andersson var. purpurascens (Andersson) Matsum., Rosa lucieae Franch. & Rochebr. ex Crép., and Vicia cracca L., whereas alien or cultivated plants consisted of a total of 16 taxa, Chloris virgata Sw., Cirsium vulgare (Savi) Ten., Coreopsis lanceolata L., C. tinctoria Nutt., Erigeron annuus (L.) Desf., Lepidium virginicum L., Medicago lupulina L., M. polymorpha L., Melilotus suaveolens Ledeb., Oenothera biennis L., Papaver rhoeas L., Symphyotrichum pilosum (Willd.) G. L. Nesom, Tamarix chinensis Lour., Trifolium pratense L., T. repens L., Veronica persica Poir.
Chaenorhinum minus, originating from Europe and North Africa, has been introduced into North America and East Asia (
Widrlechner, 1983;
Arnold 1991;
Güemes et al., 2016;
Zare et al., 2018;
POWO, 2024). In the United States, there was a case speculated to have spread along railroads after its initial introduction by a ship (
Widrlechner, 1983). Considering the surrounding environment, it is postulated, based on this case and other reports on the invasion of alien plants in the Republic of Korea, that
C. minus was unintentionally introduced to Dangjin-si by transportation vehicles and materials traveling to and from the aforementioned port during road construction in the vacant lot (
Kang et al., 2022).
Chaenorhinum minus have been introduced not only into Dangjin-si, but also into the Baekdusan Mountain (China) (
Kim et al., 2019). While currently found only in Dangjin-si in the Republic of Korea, it could spread to environments similar to its habitat and other areas via human activities. This possibility is supported by the previously mentioned case in the United States, the environmental conditions of its habitat, and the observed increase in the number of plants in the Republic of Korea.
Key for identification of genera of the tribe Antirrhineae in the Republic of Korea
1. Stem prostrate.
2. Plant usually pubescent, rooting at node; leaf hastate or sagittate ················ Kickxia Dumort. 해란초아재비속
2. Plant usually glabrous, not rooting at node; leaf reniform or orbicular ··············· Cymbalaria Hill 덩굴해란초속
1. Stem erect.