Introduction
Since Soltis et al. (1996) reported polyphyly of Saxifraga L. sensu lato, many species within the genus have been transferred to other genera, especially to Micranthes Haw. (Gornall et al., 2000; Brouillet and Gornall, 2007; Elven and Murray, 2008; Akiyama et al., 2012; Tkach et al., 2015). Micranthes used to be classified as a subgenus or section under Saxifraga s.l., which includes approximately 400 species (Engler and Irmscher, 1916; Gornall, 1987); however, various molecular phylogenetic studies have confirmed repeatedly that the Micranthes clade is distinct from Saxifraga sensu stricto (Soltis et al., 2001, 2013; Xiang et al., 2012; Prieto et al., 2013; Deng et al. 2015; Tkach et al., 2015). Moreover, it was revealed that many genera (i.e., Chrysosplenium L., Astilbe Buch.-Ham. ex D.Don, Mukdenia Koidz., Mitella L.) have a closer phylogenetic relationship with Micranthes than with the Saxifraga s.s. clade, granting genus status of Micranthes (Soltis et al., 2001; Deng et al., 2015; Tkach et al., 2015).
The segregation of Micranthes from Saxifraga s.l. was also supported by several morphological features, such as absence of cauline leaves, seed coat with vertical ridges, and unitegmic ovule (Webb and Gornall, 1989; Brouillet and Elvander, 2009). As a result, new name combinations were made for most of the Northeast Asian taxa such as M. oblongifolia (Nakai) Gornall & H. Ohba (= S. oblongifolia Nakai), M. laciniata (Nakai & Takeda ex Nakai) S. Akiyama & H. Ohba (= S. laciniata Nakai & Takeda ex Nakai), and M. manchuriensis (Engl.) Gornall & H. Ohba (= Saxifraga manchuriensis (Engl.) Komarov) (Akiyama et al., 2012). Nevertheless, taxonomic treatment of S. octopetala Nakai, a closely related species to M. manchuriensis, has not yet been conducted.
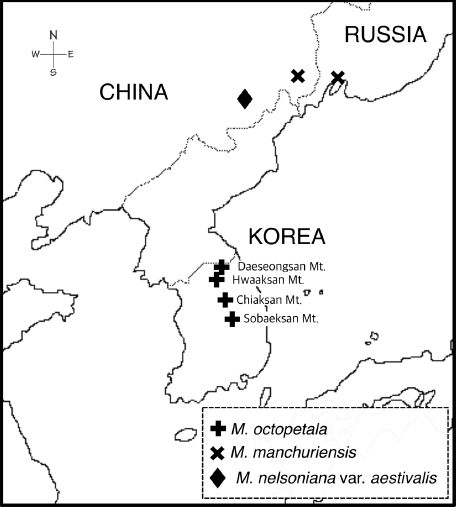
Saxifraga octopetala is a perennial herbaceous species, endemic to Korea. Nakai (1918) described the species based on four specimens collected from the eastern part of North Korea. In South Korea, it grows near small shady creeks or on moist rock crevices in high mountain areas, with the southern limit in Sobaeksan Mt. Morphologically, S. octopetala strongly resembles M. manchuriensis, which is distributed in eastern Jilin in China and Southern Primorsky Krai in Far East Russia. In the description of S. octopetala, Nakai (1918) listed the following diagnostic characters (compared with M. manchuriensis): rhizome is not fasciculate, underground stolons present, flowers arranged more densely in panicle, number of petals is not five but eight, lengths of petals and filaments are two and three times longer than sepals, respectively, and the shape of anther is spheric (vs. oblong). However, these diagnostic characters, especially the floral ones, are known to be unuseful because of disagreement between the species description and field observation. Thus sometimes S. octopetala is considered conspecific to M. manchuriensis (Chang et al., 2014).
This study was conducted to ascertain the phylogenetic position and species delimitation of S. octopetala within Micranthes. To achieve these goals, we performed phylogenetic analysis of nrITS (Internal Transcribed Spacer regions of nuclear ribosomal DNA) sequences, which have proven to be very useful for delimiting species boundaries and elucidating phylogenetic relationships of various genera in Saxifragaceae (Okuyama and Kato, 2009; Kim and Kim, 2011; Xiang et al., 2012; Gao et al., 2015; Tkach et al., 2015). In addition, the type specimens of S. octopetala and M. manchuriensis were reexamined to check the diagnostic morphological characters of the taxa. Based on the results of the phylogenetic and morphological examinations, a new name combination (Micranthes octopetala Y.I. Kim & Y.D. Kim) for S. octopetala was proposed.
Materials and Methods
Taxon sampling
A total of 114 nrITS sequences representing 20 taxa of Micranthes and three outgroup species were included for phylogenetic analysis (Table 1). The ingroup taxa were chosen to represent eight major sections of Micranthes (Tkach et al., 2015). Three outgroup taxa (Chrysosplenium alternifolium L., Chrysosplenium valdepilosum (Ohwi) S.H. Kang & J.W. Han, and Peltoboykinia tellimoides (Maxim.) H. Hara) were selected based on previous phylogenetic studies (Xiang et al., 2012; Deng et al., 2015). Among the 114 nrITS sequences, 93 sequences were newly generated in this study: 65 accessions of M. octopetala from four populations in South Korea; 24 accessions of M. manchuriensis from Jilin, China and Vladivostok, Russia; and four accessions of M. nelsoniana var. aestivalis (Fischer & C.A. Meyer) Gornall & H. Ohba from Jilin, China (Table 3). All the leaves used for the DNA sequencing were collected in the flowering period to minimize the possibility of misidentification. Voucher specimens representing each population of the above three species were deposited at KB: M. octopetala −− Daeseongsan Mt. (KYI2013106, KYI2013107), Hwaaksan Mt. (KYI2013036, KYI2013038), Chiaksan Mt. (KYI2013128, KYI2013129), Sobaeksan Mt. (KYI2013141, KYI2013160); M. manchuriensis −− Jilin (KYI2013168, KYI2013169), Vladivostok (KYI2013223, KYI2013224); and M. nelsoniana var. aestivalis −− Jilin (KYI2013185, KYI2013202). The nrITS sequences of other Micranthes taxa and three outgroup species were downloaded from GenBank (Table 1).
DNA extraction, PCR, and phylogenetic analysis
DNA was extracted from dried leaves using DNeasy Plant Mini Kits (Qiagen, Germany) in accordance with the manufacturer's instructions. PCR amplification of the nrITS region was carried out in a total reaction volume of 50 μL containing 5 μL of Takara 10X Ex-Taq buffer, 4 μL of 2.5 mM dNTPs, 0.5 μL each of forward and reverse primers, 0.25 μL of 5 U/μL Ex-Taq polymerase, 10−20 ng of template DNA, and distilled water up to the final volume. The thermocycling profile consisted of an initial denaturation step at 95oC for 5 min; followed by 30 cycles of 1 min at 95oC, 1 min at 57oC, and 1 min at 72oC; and a final extension step of 10 min at 72oC. The products were purified with a QIAqiuck PCR purification kit (Qiagen, Germany) according to the manufacturer's instructions. Purified double-stranded PCR products were used for determining the DNA sequence of the nrITS region using the automatic DNA sequencer ABI PRISM 377 (PE Applied Biosystems). The nrITS regions were amplified and sequenced using primers designed by White et al. (1990), except for ITS1, which differed by the two italicized bases (5-GGA AGG AGA AGT CGT AAC AAG G-3).
DNA sequences were aligned with MAFFT (Katoh et al., 2002). The alignment was further examined and slightly edited manually as needed. Gaps introduced from the alignment were treated as missing characters in subsequent analyses. Phylogenetic analyses were performed using Maximum Parsimony (MP), Maximum Likelihood (ML), and Bayesian inference (BI) methods. MP and ML analyses were conducted in PAUP* ver.4.0b10 (Swofford, 2002). BI was conducted with MrBayes ver. 3.1.2 (Ronquist and Huelsenbeck, 2003). For the MP and ML phlyogenetic analyses, heuristic searches with the MULTREES option, ten random entries of taxa, and TBR (Tree Bisection-Reconnection) branch swapping were selected. TVM + G model was applied for ML and BI analyses as the best fit evolutionary model, which was calculated by using jModelTest 2.1.7 (Darriba et al., 2012). Bootstrap analyses (Felsenstein, 1985) with 1000 replicates were conducted to evaluate the degree of support for given clades for ML and MP analysis using the same options as tree search. In BI analysis, each Markov chain was started from a random tree and run for 2,000,000 generations, sampling trees every 200 generations, with four independent chains running simultaneously. After discarding burn-in samples (initial 2500 trees), the remaining samples were retained for the construction of a final tree with posterior probabilities for given clades. The strict consensus tree of 24 equally parsimonious trees (Fig. 2) and a BI tree (Fig. 3) were presented for the discussion of phylogenetic relationships.
Morphology
Specimens of Micranthes nelsoniana var. aestivalis, M. manchuriensis, and M. octopetala collected by the authors as well as type specimens of M. octopetala (= S. octotpetala) deposited in TI and M. manchuriensis were used to verify the diagnostic characters of M. octopetala. Two type specimens of M. manchuriensis (= Saxifraga manchuriensis), which are assumed to have been referred by Nakai (1918), were observed using the high resolution digital images provided by K (http://apps.kew.org/herbcat/getImage.do?imageBarcode=K000618434) and HUH (http://kiki.huh.harvard.edu/databases/specimen_search.php?mode=details&id=255501). The image of the specimen (isotype) provided by HUH for the discussion of taxonomic identity of S. octopetala is presented in Fig. 4.
Results
Sequence variation and ribotypes
The ribotypes (nrITS sequence types) found in each population of M. octopetala, M. manchuriensis, and M. nelsoniana var. aestivalis are listed in Table 2. All 65 accessions in M. octopetala have nrITS sequence size of 664 bp. They were clustered into 6 ribotypes according to variation observed in the 3 variable nucleotide sites. Among the four populations, Chiaksan Mt. exhibited the highest diversity in nrITS sequence types (Table 3). The sequences of 24 accessions of M. manchuriensis were 657 to 658 bp in length and were grouped into 5 ribotypes based on variations in 4 nucleotide sites (Table 2). In M. manchuriensis, higher ribotype diversity was observed in the Vladivostok population (Table 3). The sequence lengths of M. nelsoniana var. aestivalis were all 659 bp. They were classified into 2 ribotypes by the variation in 3 nucleotide sites: type 1 and 2 with three and one accession, respectively (Table 3).
The sequence lengths of the nrITS regions of the examined taxa ranged from 637 bp (Micranthes brachypetala (Malyschev) Gornall & H. Ohba) to 669 bp (Peltoboykinia tellimoides (Maxim.) H. Hara). The aligned nrITS region was 789 bp long including 358 variable sites, among which 272 sites were parsimony informative. M. octopetala, M. manchuriensis, and M. nelsoniana var. aestivalis showed a total of 49 sites of sequence variation, which were useful in distinguishing each taxon. The amount of nrITS sequence difference between M. octopetala and M. manchuriensis ranged from 18 to 26 bp, depending on accessions (or their ribotypes) compared.
Phylogenetic analyses
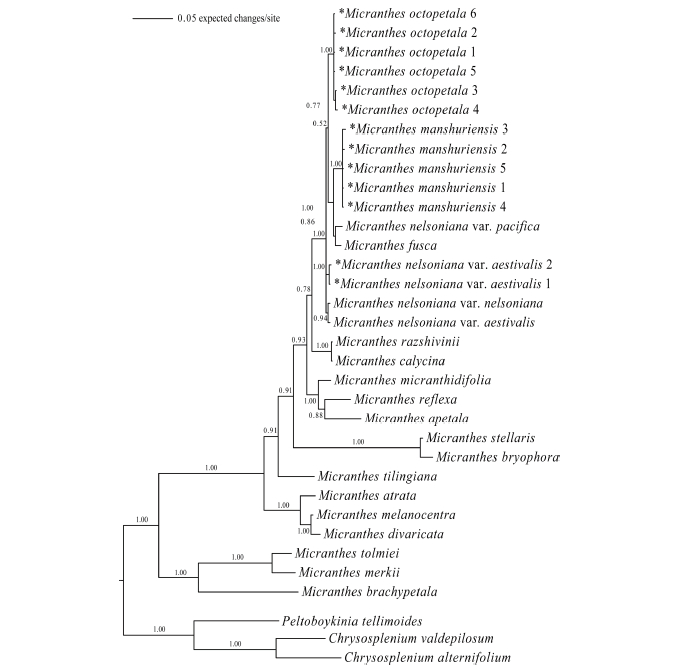
Twenty-four equally parsimonious trees were obtained as a result of MP analysis. The length of the phylogenetic tree was 806, and CI (Consistency Index) and RI (Retention Index) were 0.6725 and 0.7604, respectively. The strict consensus tree is shown in Fig. 2. The BI tree (Fig. 3) exhibited almost identical topology to the strict consensus tree. In both trees, the Micranthes clade was evident with high bootstrap values (98/ 100 in MP/ML analysis) and posterior probability (1.00). S. octopetala grouped with M. manchuriensis, M. nelsoniana, and M. fusca within the section Rotundifoliatae (A.M. Johnson) Tkach of the Micranthes clade, which was supported by bootstrap values (97/79 in MP/ML analysis) and posterior probability (1.00).
Sixty-five accessions of S. octopetala formed a robust clade with high bootstrap values (100/96 in MP/ML analysis) and posterior probability (1.00). All 24 accessions of M. manchuriensis collected from China and Russia also formed a strong monophyletic group with high bootstrap values (100/96 in MP/ML analysis) and posterior probability (1.00). In both trees, M. manchuriensis showed a sister group relationship with a clade consisting of M. nelsoniana var. pacifica and M. fusca (Maxim.) S. Akiyama & H. Ohba (bootstrap values of 78/81 in MP/ML analysis; posterior probability of 1.00), making the M. nelsoniana complex non-monophyletic.
Morphological examination
As a result of examining four type specimens of Saxifraga octopetala, scanned images of Micranthes manchuriensis types, and specimens collected by the authors from 6 populations in Korea, China, and Russia, no definite morphological differences in the leaf shape or floral features were found between the two taxa. Careful observation on type materials and other specimens revealed that both S. octopetala and M. manchuriensis have flowers with 8 (or sometimes 9) petals. It was confirmed that the type specimen of M. manchuriensis deposited in HUH was a complex collection consisting of a M. manchuriensis individual and an inflorescence assumed to be from another species of Micranthes, probably M. nelsoniana var. aestivalis, which has a loose inflorescence and flowers with 5 petals (Fig. 4). The presence of underground stolons, a diagnostic character state of S. octopetala suggested by Nakai (1918), was not clearly seen in the type specimens although they were observed from the specimens collected in this study.
Discussion
Delimiting or recognizing species by using DNA data and subsequent reexamination of non-molecular traits is an active area of systematic research (Lpez et al., 2007; Wiens, 2007; Okuyama and Kato, 2009; Ruiz-Sanchez and Sosa, 2010; Efimov, 2013; Boatwright et al., 2015). The nrITS sequence data utilized for this study proved to be very useful in delimiting the Korean endemic Saxifraga octopetala (= Micranthes octopetala) from M. manchuriensis and other related taxa such as M. nelsoniana var. aestivalis (Table 2). Considering the strong morphological similarities between S. octopetala and M. manchuriensis, an nrITS sequence difference of 18 to 26 bp between the two taxa is a noteworthy result. This suggests that substantial genetic divergence has occurred between these lineages. The resulting nrITS trees (Figs. 2, 3) also indicate that S. octopetala is a well segregated lineage that merits a distinct species status within Micranthes. Moreover, merging S. octopetala with M. manchuriensis is phylogenetically inappropriate as the latter species shows a closer relationship with M. nelsoniana var. pacifica and M. fusca than with S. octopetala (Figs. 2, 3).
Although all other species of Saxifraga s.l. distributed in Korea have been transferred to Micranthes (Akiyama et al., 2012), the taxonomic treatment of S. octopetala has been neglected, probably because of its limited distribution and ambiguous species entity, especially in relation to the unclear species delimitation between the species and M. manchuriensis. The morphological difference between the two species was not apparent because most of the diagnostic characters of S. octopetala (especially reproductive ones) proposed by Nakai (1918) were mistaken, making them inapplicable for species identification. For example, both S. octopetala and M. manchuriensis were confirmed to have 8 (sometimes 9) petals, yet Nakai (1918) stated that the latter species has 5 petals.
The reason why Nakai (1918) recognized the petal number of M. manchuriensis as 5 and not 8 needs to be considered. By paying attention to the mis-mounted inflorescence of other species (probably, M. nelsoniana var. aestivalis; Fig. 4), Nakai (1918) seems to have misinterpreted the floral characteristics of M. manchuriensis. Neither Engler (1872) nor Komarov (1903) mentioned the number of petals in their description or taxonomic treatment of M. manchuriensis. It is highly probable that Nakai (1918) determined the floral features (number of petals, stamen characteristics, etc.) of M. manchuriensis by examining the mis-mounted loose inflorescence, which was relatively easier for observation of flowers, rather than the inflorescence of the genuine individual with congested flowers. Therefore, it can be inferred that the diagnostic characters of the floral part listed in Nakai (1918) are probably those of M. nelsoniana var. aestivalis.
It is intriguing that M. manchuriensis showed a closer phylogenetic relationship with M. nelsoniana var. pacifica and M. fusca than with S. octopetala, which sometimes is considered conspecific to M. manchuriensis (Chang et al., 2014). It should be emphasized that this result is based on examination of multiple (up to 65) accessions representing each taxon and on careful collection of leaves (from flowering individuals) for DNA analysis. Identical topology and relatively high statistical supports for the relationship (Figs. 2, 3) suggest that the discordance between phylogenetic affinity and morphological similarity is largely irrelevant to the employed phylogenetic analysis methods.
Symplesiomorphy of M. manchuriensis and S. octopetala might be the cause of the morphological similarity. In other words, morphological traits of their ancestor seem to have continued to exist in both species even after genetic differentiation, leading to the apparent morphological similarity. The evolutionary phenomenon known as morphological stasis (Larson, 1989) has been reported in several plant groups including Aralia sect. Dimorphanthus (Wen, 2000), Liquidambar (Hoey and Parks 1991; Shi et al., 1998), Magnolia sect. Rytidospermum (Qiu et al., 1995), Osmorhiza (Wen et al., 2002), and Phryma (Nie et al., 2006). Unlike the above cases, which dealt with morphological stasis of taxa showing intercontinental disjunctive distribution, our result is noteworthy as it is observed from taxa distributed adjacent to each other in northeast Asia.
The sister group relationship between M. manchuriensis and the clade consisting of M. nelsoniana var. pacifica and M. fusca poses another taxonomic issue: non-monophyly of M. nelsoniana. It is known that many intraspecific taxa of M. nelsoniana are widely distributed in cool-temperate regions in the northern hemisphere (Brouillet and Elvander, 2009). Non-monophyly of the species complex was also observed in a previous study (Tkach et al., 2015). As with other species complexes, many of the intraspecific taxa have often been treated as distinct species. For example, M. nelsoniana var. pacifica has been treated as Saxifraga pacifica (Hultdèn) Zhmylev & V.V. Petrovsky. Thus, the current taxonomy of the M. nelsoniana complex needs to be tested by more detailed phylogenetic and morphological reexamination using extensive sampling encompassing all known taxa in the species complex in the northern hemisphere. Sufficient taxon sampling will be important because there are reports that the relationship of ingroups often shows different patterns if there is an insufficient number of taxonomic groups representing various lineages in the phylogenetic analysis (Zwickl and Hillis, 2002; Jockusch et al., 2015).
Meanwhile, the presence or absence of underground stolons would be a good character to distinguish S. octopetala and M. manchuriensis. Also, the rhizome of M. manchuriensis tends to be thicker than that of S. octopetala, although this feature is less consistent than the feature of underground stolons. Our study suggests that vegetative characters, including underground parts, are more useful than the known floral ones in morphological delimitation of these taxa. The characteristics of vegetative parts, such as growth pattern of sterile branches, have been verified as important in recognizing new cryptic species in the neighboring genus Chrysosplenium (Han et al., 2011; Kim and Kim, 2014; Kim 2015). The morphological difference between the two taxa should be explored further through a detailed comparative study of additional characters such as fruit and seed morphology, which have not been examined thus far. Based on the result of molecular phylogenetic analysis and morphological difference (i.e., presence or absence of underground stolons) between S. octopetala and M. manchuriensis, a new combination for S. octopetala is presented below.
Taxonomic Treatment
Micranthes octopetala (Nakai) Y. I. Kim & Y. D. Kim, comb. nov.
Basionym: Saxifraga octopetala Nakai, Bot. Mag. (Tokyo) 32: 230. 1918. TYPE: Korea. Chagang-do, Taeheungri− Sanyang, 21 July 1914 T. Nakai 6439 (lectotype, TI!); Hamgyong-bukto, Ranan, 1913, I. Ono s.n. (syntype, TI!); Kangwon-do, Ouensan, July 1906, U.J. Faurie 377 (syntype, TI!); Rason, Solsinsan, 28 August 1914, T. Nakai 6450 (syntype, TI!).
Herbs, perennial, to 15−30 cm, with underground stolons; rhizomes short, transversely elongating. Flowering stems erect, leafless, glandular, villous. Leaves 1−4, basal; stipules absent; petiole 4.7−13 cm long, sparsely glandular piliferous distally; blade reniform to orbicular-cordate, 3.8−8.5 cm long, 4.2− 10.3 cm wide, base reniform, apex rounded, margin 23−32- crenate, abaxially and marginally strigulose, palmately veined. Inflorescence paniculate, compact, many flowered, 3−9.3 cm long; bracts lanceolate or linear, 4−7 mm long; pedicels glandular hairy. Flower: bisexual, radially symmetric; sepals 8 (rarely 9), reflexed, sublanceolate, 2.0−2.5 mm long, 0.4− 0.7 mm wide, apex subobtuse, glandular hairy on abaxial surface, 1-veined, margin glandular ciliate; petals 8 (rarely 9), white, oblong-oblanceolate, ca. 4 mm long, ca. 1 mm wide, apex subobtuse or rounded or minutely retuse, 1-veined; stamens 15−17, 3.6−5.8 mm long; filaments white, slightly clavate; anthers spheric to oblong, orange; carpels 2 (or 3), superior, united only at base, white, conical; styles 2 (or 3), erect or slightly curved, 3.3−3.4 mm long; stigma capitate. Capsules erect, divergent, 4.5−6.0 mm long.
Korean Name: Gu-sil-ba-wi-chui (구실바위취)
Flowering: Jun. to Jul.
Distribution: Korea (endemic).
Korea: N Korea (HB, HN), GG, GW, CB. Wet places along shady creeks or on rock crevices in high mountain areas.