








Artemisia L. is the biggest genus of tribe Anthemideae of Asteraceae (Bremer and Humphries, 1993; Heywood and Humphries, 1997), comprising about 500 taxa. Many Artemisia species are distributed widely on temperate region and the middle-high latitude of the Northern Hemisphere. However, some species are distributed in arid region and the Southern Hemisphere (McArthur, 1979; Mabberley, 1990; Ling, 1991, 1992, 1995a, 1995b; Bremer and Humphries, 1993).
The scientific name, taxonomic position, and geographical distribution of A. gmelinii Weber ex Stechm. and its allied taxa are currently confusing (Boyko, 1990). A. gmelinii was described (No. 106, 107) and illustrated (Tab. LVI. Figs. 1, 2, 3) by Gmelin (1749) without a scientific name in the Flora of Siberia at first. The scientific name of A. gmelinii was suggested by Weber and validated by Stechmann (1775). When A. gmelinii was published, the correct information described and illustrated by Gmelin (1749) was not cited. After that, Besser (1834) has suggested that No. 106, Tab. LVI, and Fig. 1 in the description and illustration of Gmelin (1749) are typification characters of A. gmelinii. He has described it as a small shrub (30–40 cm height), bipinnate divided leaves, linear and obtuse leaflets with green of adaxial, light gray of abaxial. Ledebour (1805) has published A. sacrorum and cited No. 107, Tab. LVI, and Fig. 2 of Gmelin (1749) as typification characters. According to the descriptions of Ledebour (1805), A. sacrorum was a subshrub, erect, longer than about 50 cm, branched, ridged stem with white-gray and glabrous on adaxial leaf surface, dense trichome with white-gray on abaxial leaf surface, and petiole attenuate. Later, Ledebour (1833) recognized A. sacrorum α. latiloba (dark green on adaxial leaf surface and white-gray on the abaxial leaf surface), β. intermedia (yellowish-green on adaxial leaf surface and white-gray on abaxial leaf surface), and γ. minor (=A. gmelinii). He treated A. sacorum as a synonym of β. intermedia and cited with Tab. LVI and Fig. 2. In addition, A. gmelinii was treated by him as a synonym of γ. minor. He cited Tab. LVI and Fig. 1 as the typification of A. gmelinii, which has caused confusion. Besser (1834) recognized A. gmelinii under α. biebersteiniana, β. legitima, and γ. gebleriana. He published A. messerschmidtiana as a new species within the following two intraspecific taxa: α. viridis, and β. incana. Since α. viridis has dense trichome only on abaxial leaf, he treated A. sacrorum as a synonym based on Gmel. Fl. Sib. no. 107. Fig. 2. However, β. incana has dense trichome with white-gray on both surfaces. He has insisted that A. messerschmidtiana is only distributed in east Siberia, α. viridis is distributed in Dahuria, and β. incana is distributed in Baikal. The taxonomic status and scientific names of Artemisia gmelinii and its allied taxa have long been debated. There is still confusion now (Komarov, 1907; Nakai, 1911, 1912; Pampanini, 1927; Kitamura, 1937; Poljakov, 1961; Ling, 1988; Lee, 1996).
Thus, the objective of this study was to reevaluate the taxonomic status of A. sacrorum and its allied taxa based on morphological, cytological (Park et al., 2009), and palynological studies (Park and Chung, 2012) to provide correct names for them based on original descriptions and type specimens.
Materials and Methods
Type specimens, deposited in Andong National University (ANH), Seoul National University (SNUA), Kangwon National University (KWNU), Korea National Arboretum (KH), Chungbuk National University (CBU), Jeju National University (CNUH), Institute of Botany, Chinese Academy of Sciences (PE), Tokyo university (TI), and Kyoto university (KYO) were examined (Appendix). Materials from various populations were collected from Sep. 2006 to Oct. 2010 for this study. Qualitative and quantitative characters were observed, measured, and photographed using a stereoscopic microscope (Olympus AX-70, Japan; Olympus DP2-BSW, Germany). The main characters were also illustrated (Fig. 4). All voucher specimens were stored in ANH.
Results and Discussion
Boyko (1990) has reconsidered the taxonomic history of A. gmelinii and its allied taxa (A. sacrorum and A. messerschmidtiana). He has stated that A. gmelinii is widely distributed in Siberia but not distributed in the Far East (Russia, Korea, China, Japan, etc.). However, A. sacrorum and A. messerschmidtiana are distributed in the Far East. Its route of distribution is from the Ural Mountains to Mongol, China, Japan, and the Korean Peninsula. Maximowicz (1872) and Komarov (1907) have inferred that A. sacrorum is distributed in the Far East whereas A. gmelinii is not distributed there. After reviewing the aforementioned papers, habitats, and specimens, the 3 taxa are considered to be distributed in the Korean peninsula (Fig. 5). The taxonomic opinions are described here.
Key to the varieties of Artemisia sacrorum
1. Leaf blades sparsely pubescent on both surfaces ··············································································· var. iwayomogi
1. Leaf blades densely villous on abaxial surface
2. Leaf blades sparsely villous with white glandular on adaxial surface and densely villous on abaxial surface ································································ var. sacrorum
2. Leaf blades densely villous on both surfaces ················································································· var. incana
Artemisia sacrorum Ledeb., Mem. Acad. Imp. Sci. 5:571 (1805)
var. sacrorum (Fig. 1)
Korean name: Teol-san-ssuk (털산쑥: Lee, 1996).
A. messerschmidtiana Besser, Nouv. Mém. Soc. Imp. Naturalistes Moscou 3: 27 (1834).
A. sacrorum var. minor f. discolor Kom., Fl. Manschur. 3(2): 664 (1907).
A. gmelini var. discolor (Kom.) Nakai, Fl. Kor. 2: 31 (1911).
A. freyniana f. discolor (Kom.) Kitag., J. Jap. Bot. 41: 367 (1936).
A. sacrorum subsp. manshurica Kitam., Acta Phytotax. Geobot. 7: 66 (1938).
A. sacrorum var. messerschmidtiana (Besser) Y. R. Ling, Bull. Bot. Res., Harbin 8(4): 13 (1988).
Subshrubs. Roots thin fibrous. Rhizomes long creeping, woody. Stems 52–108 cm tall, 1.9–3.5 mm thick; indumentum silky pubescent while young, sparsely arachnoid-pubescent later. Lower leaves withering before flowering. Middle leaves petiolate 14.9–21.4 mm long; leaf blades triangular-ovate, elliptic-ovate, bipinnately divided, 43.1–65.1 mm long, 39.1–52.7 mm wide, sparsely villous pubescent with white glandular on adaxial surface, densely villous pubescent on abaxial surface; segments 5–7 pairs, lanceolate, pinnately divided, 19.1–25.3 mm long, 7.1–11.6 mm wide; ultimate segments linear-lanceolate, triangular pectinate; rachis triangular pectinate. Upper leaves small, leafy bracts, simple. Inflorescence broad panicle. Capitula long pedunculate, nodding; involucre subglobose, 2.3–2.7 mm long, 1.9–2.4 mm wide; phyllaries 4–5 seriate, laxly imbricate, densely villous; outer phyllaries ovate-lanceolate, green, narrowly membranous margin, acute at apex; middle phyllaries lanceolate, green, middle membranous margin, obtuse at apex; inner phyllaries obovate, broadly membranous margin, rounded at apex. Receptacle conical, glabrous with glandular. Marginal female florets 9–14 in number, corolla 0.8–1.1 mm long, conical with cuneate shaped glandular, 2–3 toothed irregularly at apex; style 0.8–1.3 mm long, longer than corolla; stigma 0.5–0.7 mm long, 2-divergent, somewhat emarginate at apex; ovary fertile, 0.5–0.8 mm long. Disk bisexual florets 7–11 in number, corolla 1.3–1.6 mm long, narrowly tubular without throat, cuneate shaped glandular, 5-toothed regularly at apex; style 0.8–1.3 mm long, longer than corolla; stigma 0.4–0.6 mm long, 2-divergent, combed funnelform at apex; ovary fertile, 0.5–0.8 mm long; anthers 5 in number, 2 acute basal appendages, apical appendage acute, triangular, 0.5–0.7 mm long.
Type: Russia. Tibet, Sibir (type: ?).
Flowering: Sep. to Oct.
Distribution: China, Russia, Korea, Afghanistan, Mongolia.
Distribution of Korea: Hamgyeongbuk-do, Yanggang-do, Hwanghae-do.
Habitat: Hills, waysides.
Note: A. sacrorum Ledeb var. sacrorum can be distinguished from other related taxa by small subshrub, non-glandular trichome eliminated at flowering time on adaxial leaf surface, white glandular trichome distributed on adaxial leaf surface, non-glandular trichome of white-gray on abaxial leaf surface and phyllaries (Fig. 1). It has the same taxon as A. messerschmidtiana treated as A. sacrorum var. messerschmidtiana by Ling (1988). However, A. messerschmidtiana needs to be treated as a synonym of A. sacrorum because its priority and characters with densely villous on only abaxial leaf surface. According to specimens, this taxon is restrictively distributed from Siberia to Manchuria including North Korea. It is not distributed in South Korea or Japan (Fig. 5). Kitamura (1937) has published A. sacarorum subsp. manshurica with the size of capitula, the shape of a leaf, and geographical delimitation by comparing to A. sacrorum of Altai region and to the illustration (Icones Pl. fl. Ross. tab. 471) of Ledebour (1834) without considering the geographical variation through observation of limited specimens. In Korea, the scientific name of A. freyniana f. discolor combined by Kitagawa (1996) is used. However, this scientific name was published by Komarov (1907) without considering geographical variations. Therefore, we treated those names as synonyms of A. sacororum including A. gmelini var. discolor by Nakai (1911).
Artemisia sacrorum Ledeb. var. incana (Besser) Y. R. Ling, Bull. Bot. Res., Harbin 8(4): 13 (1988). (Fig. 2)
Korean name: Huin-san-ssuk (흰산쑥: Lee, 1996).
A. messerschmidtiana var. incana Besser, Nouv. Mém. Soc. Imp. Naturalistes Moscou 3: 27 (1834).
A. sacrorum f. vestita Kom., Fl. Manschur. 3(2): 664 (1907).
A. gmelini var. vestita (Kom.) Nakai, Fl. Kor. 2: 31 (1911).
A. freyniana f. vestita (Kom.) Kitag., J. Jap. Bot. 41: 367 (1936).
A. sacarorum var. vestita Kitam., Acta Phytotax. Geobot. 7: 66 (1938).
Subshrubs. Roots thin fibrous. Rhizomes long creeping woody. Stems 56–105 cm tall, 1.5–3.9 mm thick; indumentum silky pubescent while young, sparsely arachnoid-pubescent later. Lower leaves withering before flowering. Middle leaves petiolate 13.1–19.1 mm long; leaf blades triangular-ovate, elliptic-ovate, bipinnately divided, 41.6–61.4 mm long, 41.1–59.1 mm wide, densely villous pubescent with glandular on both surfaces; segments 5–7 pairs, lanceolate, pinnately divided, 21.9–28.5 mm long, 8.2–11.2 mm wide; ultimate segments linear-lanceolate, triangular pectinate; rachis triangular pectinate. Upper leaves small, leafy bracts, simple. Inflorescence broad panicle. Capitula long pedunculate, nodding; involucre subglobose, 2.7–3.0 mm long, 1.9–2.7 mm wide; phyllaries 4–5 seriate, laxly imbricate, densely villous with glandular; outer phyllaries ovate-lanceolate, narrowly membranous margin, acute at apex; middle phyllaries lanceolate, middle membranous margin, obtuse at apex; inner phyllaries obovate, broadly membranous margin, rounded at apex. Receptacle conical, glabrous with glandular. Marginal female florets 5–10 in number, corolla 0.6–1.2 mm long, conical with cuneate shaped glandular, 2–3 toothed irregularly at apex; style 0.7–1.3 mm long, longer than corolla; stigma 0.5–0.8 mm long, 2-divergent, somewhat emarginate at apex; ovary fertile, 0.5–0.9 mm long. Disk bisexual florets 8–13 in number, corolla 1.2–1.7 mm long, narrowly tubular without throat, cuneate shaped glandular, 5-toothed regularly at apex; style 0.6–1.2 mm long, longer than corolla; stigma 0.3–0.5 mm long, 2-divergent, combed funnel form at apex; ovary fertile, 0.5–0.8 mm long; anthers 5 in number, 2 acute basal appendages, apical appendage acute, triangular, 0.4–0.6 mm long.
Type: Sibir (type: ?).
Flowering: Sep. to Oct.
Distribution: China, Korea, Mongolia.
Distribution of Korea: Hamgyeongbuk-do, Yanggang-do.
Habitat: Hills, waysides, shrublands, slopes, often dominant on S slopes, roadsides, forest steppes.
Note: This taxon was recognized as a new species of A. messerschmidtiana by Besser (1834) at first with two infraspecific taxa: α. viridis, β. incana. He treated A. sacrorum as a synonym of α. viridis (= A. sacrorum var. sacrorum) characterized by glabrous adaxial leaf surface and white-gray trichome on abaxial leaf surface. Also, β. incana was first described with characters of white-gray trichome on both leaf surfaces. After that, Ling (1988) classified α. viridis and β. incana as varieties of A. sacrorum because of their morphological similarities. We also found out that there was no difference among them except the distribution pattern of trichome between A. sacrorum and α. viridis, β. incana. This taxon is considered to have geographical distribution from Siberia to Manchuria, including North Korea. However, it is not distributed in South Korea or Japan (Fig. 5). In Korea, the scientific name of A. freyniana f. vestita combined by Kitagawa (1996) is used. However, the name of A. sacrorum var. minor f. vestita published by Komarov (1907) is a combination without considering geographical variations. We treated A. freyniana f. vestita as a synonym of A. sacororum var. incana including A. gmelini var. vestita suggested by Nakai (1911) and A. sacarorum subsp. manshurica var. vestit used by Kitamura (1937).
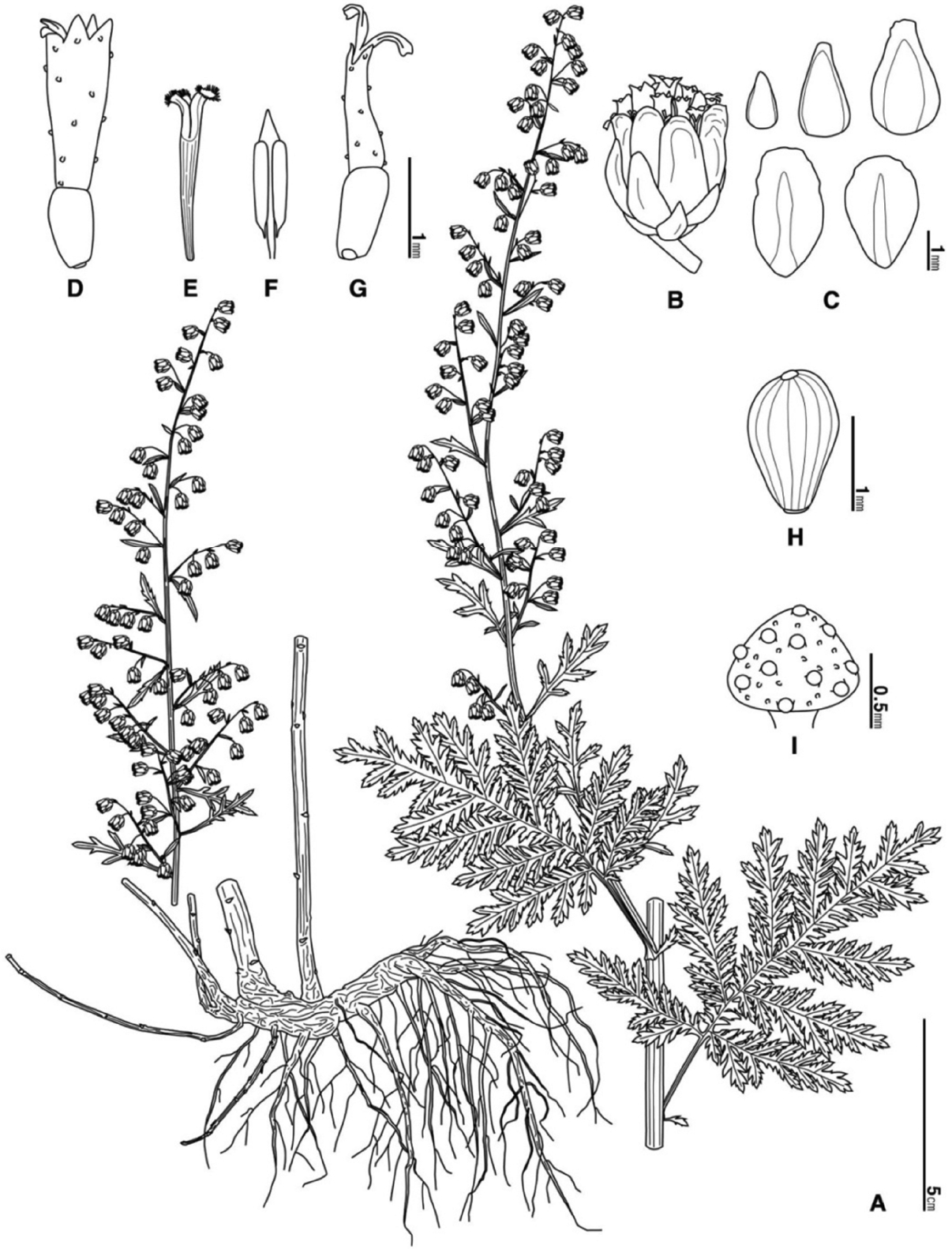
Artemisia sacrorum Ledeb. var. iwayomogi (Kitam.) M. S. Park & G. Y. Chung, comb. et stat. nov. (Figs. 3–4).
Korean name: Deo-wi-ji-gi (더위지기: Lee, 1996).
Basionym: A. iwayomogi Kitam., Acta Phytotax. Geobot. 7: 65 (1938).
A. sacrorum subsp. iwayomogi (Kitam.) Vorosch., Byull. Moskovsk. Obshch. Isp. Prir., Otd. Biol. 102(1): 68 (1997).
A. messerschmidtiana f. laxiflora Nakai, 2: 31 (1911).
A. sacrorum subsp. laxiflora (Nakai) Kitag., Neo-Lineam. Fl. Mansh. 431 (1979).
Subshrubs. Roots thin fibrous. Rhizomes long creeping woody. Stems 71–153 cm tall, 2.8–4.2 mm thick; indumentum silky pubescent while young, sparsely arachnoid-pubescent later. Lower leaves withering before flowering. Middle leaves petiolate 13.3–20.3 mm long; leaf blades triangular-ovate, elliptic-ovate, bipinnately divided, 53.7–105.4 mm long, 40.8–67.2 mm wide, sparsely villous with glandular on both surfaces; segments 5–8 pairs, lanceolate, pinnately divided, 24.5–35.8 mm long, 8.3–12.4 mm wide; ultimate segments linear-lanceolate, triangular pectinate; rachis triangular pectinate. Upper leaves small, leafy bracts, simple. Inflorescence broad panicle. Capitula long pedunculate, nodding; involucre subglobose, 2.7–3.1 mm long, 2.7–3.0 mm wide; phyllaries 4–5 seriate, laxly imbricate, sparsely villous with glandular; outer phyllaries ovate-lanceolate, narrowly membranous margin, acute at apex; middle phyllaries lanceolate, middle membranous margin, obtuse at apex; inner phyllaries obovate, broadly membranous margin, rounded at apex. Receptacle convex, glabrous, rarely a few T-shaped trichome with glandular. Marginal female florets 5–12 in number, corolla 0.7–1.1 mm long, conical with cuneate shaped glandular, 2–3 toothed irregularly at apex; style 0.7–1.0 mm long, longer than corolla; stigma 0.4–0.6 mm long, 2-divergent, somewhat emarginate at apex; ovary fertile, 0.5–0.8 mm long. Disk bisexual florets 5–8 in number, corolla 1.3–1.5 mm long, narrowly tubular without throat, cuneate shaped glandular, 5- toothed regularly at apex; style 0.9–1.3 mm long, longer than corolla; stigma 0.3–0.5 mm long, 2-divergent, combed funnelform at apex; ovary fertile, 0.5–0.7 mm long; anthers 5 in number, 2 acute basal appendages, apical appendage acute, triangular, 0.5–0.7 mm long. Achenes brown, pyriform, 1.2–1.5 mm long, glabrous. Chromosome number 2n = 54.
Type: Japan. Hokkaido, Prov. Shiribeshi, Setana, 13 Oct. 1935. S. Kitamura (type: in KYO!).
Flowering: Sep. to Oct.
Distribution: China, Russia, Japan, Korea, Afghanistan, N India, Kazakhstan, Mongolia, Nepal, N Pakistan.
Distribution of Korea: All provinces.
Habitat: Hills, waysides, shrublands, slopes, often dominant on slopes, roadsides, forest steppes.
Note: A. gmelinii has been used as a correct name for this taxon (Lee, 1996). After reviewing the original description and illustration of Besser (1834) and Ledebour (1805; 1833) through typification and the illustration (Gmel. Fl. Sib. No. 106, Tab. LVI. Fig. 1), A. gmelinii is widely distributed in Siberia. However, it is not distributed in the Far East (Maximowicz; 1872, Komarov; 1907; Boyko, 1990). Kitamura (1937) has published A. iwayomogi (nomen novum) instead of a wrong scientific name that some scholars have misused. It was distinguished from A. sacrorum of Altia region. He mentioned that A. sacrorum had small sized plant and leaf, acute leaflet, and definitely different geographical distribution. He described that A. iwayomogi was distributed in Japan, Sakhalin, Kuril Islands, the Korean peninsula, and the Far East of Manchuria. After observing specimens of Russia, China, and Japan, A. gmelinii is mainly distributed in Siberia but not the Far East, while A. sacrorum var. sacrorum (=A. messerschmidtiana) and A. sacrorum var. incana have expanded their distribution from Ural mountains to Dahuria of the northeast, Far East, the northeast China, and North Korea. In addition, A. sacrorum var. iwayomogi is distributed in Mongolia, northeast China, Japan, Sakhalin, Kuril Islands, the Far East of Manchuria, and the entire Korean peninsula including the area mentioned by Kitamura (1937). According to the original description and type specimen of A. iwayomogi published by Kitamura (1937), the typification of A. sacrorum var. iwayomogi distributed in South Korea is the same. The distribution of A. sacrorum var. iwayomogi almost overlaps with A. sacrorum var. sacrorum and A. sacrorum var. incana. Considering that there is no difference except that A. iwayomogi has no non-glandular trichome on both leaf surface or phyllaries of capitula at flowering time, A. iwayomogi needs to be treated as a variety of A. sacrorum.
The chromosome number of A. sacrorum var. sacrorum and A. sacrorum var. incana has been reported to be 2n = 2x = 18 (Kaul and Bakshi, 1984; Volkova and Boyko, 1985). The reported chromosome number of A. sacrorum var. iwayomogi is 2n = 6x = 54 (Park et al., 2009). Inferring evolution trend from geographical distribution, morphological characters, and chromosome numbers of the A. sacororum group, when A. sacrorum var. sacrorum and A. sacrorum var. incana (diploid) with similar geographical distribution moved to the Far East, the Korean Peninsula, and Japan, the trichome might have been eliminated from both leaf surfaces to adapt to the environment. At that time, A. sacrorum var. iwayomogi (haxaploid) might have been divided (Fig. 5). Such hypothesis about their relationship has been supported by Park and Chung (2012). They have reported that the pattern of pollen surface, A. sacrorum var. sacrorum, and A. sacrorum var. incana has a connected base of spinule on pollen. However, A. sacrorum var. iwayomogi has a disconnected base of spinule on pollen.
Consequently, there are three varieties under A. sacrorum in the Korean peninsula: A. sacrorum var. sacrorum with white-gray non-glandular trichome on adaxial leaf surface, A. sacrorum var. incana with white-gray non-glandular trichome on both leaf surfaces, and A. sacrorum var. iwayomogi with almost glabrous on both leaf surfaces. A. sacrorum var. sacrorum and A. sacrorum var. incana are distributed mainly in Siberia, expanding from Ural mountains to Dahuria of the northeast, Far East of Russia, northeast China, and North Korea, while A. sacrorum var. iwayomogi is widely distributed in Mongolia, northeast China, Japan, Sakhalin, Kuril Islands, the Far East of Manchuria, and the Korean Peninsula.




 Artemisia gmelinii (Asteraceae)
Artemisia gmelinii (Asteraceae)  .
.  . 110-115.
. 110-115. 

