국내 벚나무류(Prunus subg. Cerasus) 식재 현황: 분당중앙공원 일대 사례연구
적 요
벚나무류 (Prunus subg. Cerasus)는 가로수 및 조경수로 전국에 가장 많이 식재하지만 외형으로 식별이 어려워서 종별 수량은 관리되지 않고 있다. 다양한 벚나무류가 식재된 성남시 분당구 분당중앙공원과 인접 지역의 식재유형과 종 구성 등의 실태를 파악하고, 조경수목 식재 관리 필요성을 논의하고자 하였다. 벚나무류의 개화기인 2021년 4월에 성남시 분당구 일대 약 6 km2 내에 식재된 벚나무류 총 5,866주를 조사하였다. 이 중 5,744주는 11분류군으로 구분하였으며, 나머지 122주는 복합적인 외부형태 특징을 보였다. 소메이요시노벚나무(Prunus ×yedoensis Matsum.)가 가장 높은 비중(52.1%)을 차지하였고, 잔털벚나무(P. serrulata Lindl. var. pubescens (Makino) Nakai), 벚나무(P. jamasakura Siebold ex Koidz.) 등의 순으로 나타났다. 제주도에 자생하는 왕벚나무(P. ×nudiflora (Koehne) Koidz.)는 본 조사에서 확인되지 않았다. 미소한 특징으로 구분되는 벚나무류의 식별을 돕고자 주요 조경용 벚나무류 11분류군에 대한 검색표를 제시하였다. 벚나무류는 자연 상태에서도 빈번하게 종간 잡종을 형성하는 것으로 알려져 있다. 외래 벚나무류와의 잡종화와 이에 따른 유전자 이입에 의해 자생종의 유전적 고유성이 손실되는 것을 막기 위해서는 자생지 주변의 식재 수종 관리와 원산지 추적이 필요하다.
주요어: 벚나무속, 벚나무아속, 소메이요시노벚나무, 왕벚나무, 잡종화
Abstract
Prunus subg. Cerasus is the most planted street and landscaping tree in South Korea, but it is difficult to identify species according to their macro-morphologies, leading to problems when attempting properly to manage species quantities. The purpose of this study is to understand the current status of plant types and species compositions in Bundang Central Park in Bundang-gu of Seongnam City and to discuss the necessity of the management of landscaping tree planting. In April of 2021, during the cherry blossom season, a total of 5,866 planted cherry trees were investigated within an area of 6 km2 of Bundang-gu in Seongnam City. As a result, 5,744 trees were sorted into eleven taxa, but the remaining 122 trees were not determined due to their complex morphologies. Prunus ×yedoensis Matsum. accounted for the highest proportion (52.1%), followed by P. serrulata Lindl. var. pubescens Nakai and P. jamasakura Siebold ex Koidz. P. ×nudiflora (Koehne) Koidz., a plant native to Jejudo Island, was not found in this survey. In order to help identify cherry trees based on micro-morphologies, an identification key was presented for the eleven taxa planted as major landscaping trees. It is known that cherry trees frequently form interspecific hybrids in nature. In order to prevent a loss of the genetic originality of native species due to hybridization and gene introgression from foreign cherry trees, it is necessary to manage planting species near the habitats of native taxa and track their origins.
Keywords: Cerasus, hybridization, Prunus, Prunus ×nudiflora, Prunus ×yedoensis
서론
벚나무속( Prunus L.)은 장미과(Rosaceae) 벚나무아과(Amygdaloideae)에 속하고 북반구 온대지역 및 아시아와 아메리카대륙의 열대지역을 중심으로 분포하는 관목 또는 교목으로서 세계적으로는 200여 종이 알려져 있다( Kalkman, 2004). 국내에는 벚나무속 식물로 21분류군이 알려져 있으며 이 중 자생종은 14분류군이다( Lee and Kim, 2007). 벚나무속은 화서와 열매 등 형태적 특징에 따라 여러 개의 독립된 속이나 아속으로 세분하기도 하였다(Linnaeus, 1753; Rehder, 1940; Kalkman, 2004). 150여 종을 포함하는 벚나무아속은 독립된 속으로 다루기도 하였지만, 최근 분자계통학적 증거에 의해 강하게 지지되는 3개의 아속(subg. Cerasus, Prunus, Padus)으로 구분한다(Shi et al., 2013; Katsuki and Iketani, 2016). 벚나무아속은 열매를 식용하는 체리류와 조경용으로 식재되는 종 대부분을 포함하며, 관상 가치가 높아서 전 세계에 널리 식재되는 소메이요시노벚나무( Prunus ×yedoensis Matsum.)도 이에 속한다(Shi et al., 2013). 산림청의 2019년 가로수 조성 실적에 따르면, 국내에 식재된 가로수 825만 주 중 벚나무류는 154만 주로 19%를 차지하며, 동 기관의 2020년 임산물 생산 현황에 의하면 매년 15만 주의 묘목이 생산된다(Korea Forest Service, 2020, 2021). 발아율이 낮아 실생묘 생산이 어려운 수종의 증식과 일관된 형질을 지닌 묘목 생산을 위해서 주로 접목을 이용하고 있으며, 대목으로는 산벚나무와 벚나무를 이용하는 것으로 알려져 있다( Kim, 2004). 다양한 벚나무류가 가로수로 식재되어 있으나 ‘왕벚나무’와 벚나무만 집계되고 있고, 대목으로 사용되는 산벚나무와 벚나무의 실체는 불분명한 실정이다. 벚나무류는 자가불화합성이 강하여 종간 잡종을 비교적 쉽게 형성하며, 품종 개량을 위해 인공잡종 형성을 유도하기도 한다( Kuitert and Peterse, 1999; Liu et al., 2007; Cho and Kim, 2019). 잡종화에 의한 형성된 개체는 결실률과 발아율이 낮아 유성생식에 의한 번식은 어렵지만 드물게 야생화되는 사례가 보고되었다( Cho et al., 2017). 국내 종묘상에 유통되는 벚나무류 외래품종은 50여 종류이나, 도입시기 등이 불명확하고, 같은 품종도 다른 명칭으로 판매하는 경우가 많다( Korea National Aboretum, 2022; Milim Botanic Garden, 2022). 이러한 외래 벚나무류의 식재는 인근 야생 개체군에 유전자 이입을 초래하여 고유종 보전에 실질적인 위협이 되는 것으로 보고되어 국내에서의 위험성을 검토할 필요가 있다(Macková et al., 2018). 벚나무류 식재 사례연구 대상지로 선정한 분당구는 성남시 전체 면적의 49.3%(69.8 km2)로, 인접한 수정구(45.5 km2), 중원구(26.4 km2)에 비해 가장 넓은 일반구이며, 1991년 분당신도시 개발이 시작된 이후 공원, 제방, 임야 등의 조성녹지 공간이 약 50%를 차지하고 있어(Seongnam City, 2020), 비슷한 시기에 식재된 가로수 및 조경수가 많을 것으로 추정되었고, 본 연구를 통해 분당 중앙공원과 인접 지역의 식재 현황을 파악하고 조경수목 식재 관리 필요성을 논의하고자 하였다.
재료 및 방법
경기도 성남시 분당구 분당중앙공원을 중심으로 서쪽의 탄천과 동쪽의 57번 지방도로를 경계로 조사구역을 설정하였고 면적은 약 6 km 2이며, 행정구역으로는 수내동, 분당동, 서현동, 정자동을 포함한다( Fig. 1). 식재 벚나무류의 식별을 위하여 개화기인 2021년 4월 5일부터 9일까지 현장조사를 하였고, 산림지역에 자생하는 벚나무류는 대상에서 제외하였다. 벚나무류를 구분하기 위해 학명과 국명은 환경부 국가생물종목록(National Institute of Biological Resources, 2021), 산림청 국가표준식물목록( Korea National Aboretum, 2022)과 기존의 연구 결과들을 참고하여 동북아생물다양성연구소 한반도관속식물목록( Northeastern Asia Biodiversity Institute, 2022)으로 정리하였고, 식재된 벚나무류의 주요 식별형질로 여겨지는 가지 습성, 엽신 털 유무, 소화경 털, 아린 털, 악통 형태, 악편 형태, 암술대 털 유무에 따라 검색표를 작성하고 이에 따라 동정하였다( Kim et al., 1998; Chang et al., 2004; Kim et al., 2005; Lee and Kim, 2007; Kim et al., 2011; Katsuki and Iketani, 2016; Jin et al., 2020).
결과 및 고찰
벚나무류 명칭의 혼용
조사지역에서 가장 높은 비율로 식재된 벚나무류는 일명 ‘왕벚나무’로 불리는 재배종인 소메이요시노벚나무( P. ×yedoensis Matsum.)이다. 해당 분류군은 1901년 동경대 교수 Matsumura에 의해 도쿄의 정원에서 재배하는 품종을 기준으로 기재되었다( Matsumura, 1901). 야생에서는 발견되지 않으며, 올벚나무( P. itosakura Siebold = P. spachiana f. ascendens)와 왜벚나무( P. speciosa (Koidz.) Nakai)의 잡종으로 밝혀진 바 있다( Katsuki and Iketani, 2016; Cho and Kim, 2019). 한편 제주도에 자생하는 왕벚나무는 프랑스 Taquet 신부가 1908년 한라산 관음사 일대에서 채집한 표본(채집번호 Taquet 4638)을 기준으로 설정된 분류군으로 P. yedoensis Matsum. var. nudiflora Koehne로 명명되었고( Koehne, 1912), 이후 종(P. nudiflora (Koehen) Koidz.)으로 승격되었다( Koidzumi, 1932). 오랫동안 ‘왕벚나무’ 원산지 논란이 있었으나, 최근 분자분류 및 유전체 연구를 통해 제주도 자생하는 왕벚나무의 모계는 올벚나무( P. itosakura Siebold = P. spachiana f. ascendens)이고 부계는 산벚나무( P. sargentii Rehder), 벚나무( P. jamasakura Siebold ex Koidz.) 또는 잔털벚나무( P. serrulata Lindl. var. pubescens (Makino) Nakai)로 확인되어 우리나라 자생 왕벚나무( P. ×nudiflora (Koehne) Koidz. = Cerasus ×nudiflora (Koehne) T. Katsuki & Iketani)는 일본산 소메이요시노벚나무와 기원이 다른 우리나라 고유 분류군으로 인정하고 있다( Cho et al., 2016; Katsuki and Iketani, 2016; Chung et al., 2017; Baek et al., 2018; Cho and Kim, 2019). 일본산 소메이요시노벚나무는 1907년 도쿄에서 수입된 3년생 묘목을 남산 왜성대공원에 심은 것을 시작으로 창경원을 비롯한 궁궐과 공원, 신작로에 식재되었다. 일제강점기를 거치면서 소메이요시노벚나무는 전국적으로 확산되었고, 벚꽃놀이는 봄철 행락문화의 대명사가 되었는데, 벚꽃이 일본을 대표하는 수종으로 인식되면서 원산지에 대한 논의가 쟁점화되었다( Kim et al., 2011). 최근 연구들을 통해 ‘왕벚나무’ 원산지 논란이 종결되었음에도 불구하고 일본산 소메이요시노벚나무와 자생 왕벚나무의 국명과 학명은 혼용되어 사용되고 있다( Jin et al., 2020). 환경부 국가생물종목록(National Institute of Biological Resources, 2021), 산림청 국가표준식물목록( Korea National Aboretum, 2022)에서는 일본산 소메이요시노벚나무의 국명을 ‘왕벚나무’로 자생 왕벚나무의 국명을 ‘제주왕벚나무’로 제시하고 있으나, 천연기념물로 지정된 왕벚나무 자생지(제156호, 제159호, 제173호)는 지정 당시 채택된 국명을 사용하고 있다( Cultural Heritage Administration, 2022). 또한 조경수로 유통되는 ‘벚나무,’ ‘산벚나무’ 등의 명칭은 야생 벚나무류를 통칭하는 단어로 사용되고 있어 혼란이 있다( Park et al., 2000; Lee et al., 2011). 우리나라 식물로서 왕벚나무를 처음 기록한 Ishidoya and Chung (1923)은 국명을 제시하지 않은 채 학명은 P. yedoensis를 사용하였으나 “한라산 상복에 자생하고, 일본에서 널리 재배하지만 자생지가 상세하지 않다”고 기록하였다. 1912년 Koehne가 발표한 제주산 P. yedoensis var. nudiflora는 연구자에 따라 P. yedoensis와 구분하여 국명을 따로 부여한 경우가 간혹 있으나( Park, 1949), 유전자를 이용한 연구가 불가능하였던 과거에는 대다수 연구자들이 일본산과 제주산 벚나무류가 서로 다른 교잡종이라는 실체를 인식하지 못했기 때문에 제주산 왕벚나무를 P. yedoensis와 동일한 종으로 이해하였다. 그 결과 P. yedoensis를 제주산 왕벚나무의 학명으로 사용하면서 국명을 ‘사꾸라( Chung et al., 1937),’ ‘왕벗나무( Park, 1949),’ ‘사구라나무( Chung, 1957),’ ‘제주벚나무( Do et al., 1958),’ ‘큰꽃벚나무(Chung, 1965)’ 등으로 다양하게 기록하였으며, Lee and Ahn (1963)이 ‘왕벗나무’를 ‘왕벚나무’로 고쳐 사용한 국명이 최근까지 사용되고 있다. 비록 실체에 대한 인식 부족으로 학명은 P. yedoensis를 사용하면서도 제주도에 자생한다는 것을 밝힘으로써 여기에 사용된 왕벚나무라는 국명은 제주산 벚나무류의 한 종인 P. ×nudiflora (Koehne) Koidz.를 지칭하는 것임을 분명히 하였다. 제주도에 자생하는 왕벚나무의 국명으로 Park (1949)이 ‘왕벗나무’를 처음으로 제시하고, Lee and Ahn (1963)이 ‘왕벚나무’로 개칭한 1963년 이후 대부분의 문헌에서 제주도산 벚나무 한 종에 대한 국명으로 왕벚나무를 사용하고 있으므로, 제주도 자생 왕벚나무를 ‘왕벚나무’로, 식재하고 있는 일본산 벚나무류를 일본 이름에서 유래한 ‘소메이요시노벚나무’로 구분하여 부를 것을 제안하며, 다음에서도 이를 적용하였다.
식재 벚나무류의 식별
조사지역 내에 식재된 벚나무류는 가지, 소지, 아린, 어린잎, 소화경, 꽃의 형태학적 식별형질을 이용하여 11분류군으로 동정하였다( Fig. 2, Table 1). 암술대 털 유무에 따라 크게 2개 그룹으로 구분되는데, 암술대에 털이 있는 그룹은 소메이요시노벚나무( Prunus ×yedoensis Matsum.), 양벚나무( P. avium (L.) L.), 처진올벚나무( P. spachiana (Lavallée ex H. Otto) Kitam.), 올벚나무( P. spachiana f. ascendens (Makino) Kitam.) 등 4분류군이며, 암술대에 털이 없는 그룹은 산벚나무( P. sargentii Rehder), 왜벚나무( P. speciosa (Koidz.) Nakai), 잔털벚나무( P. serrulata Lindl. var. pubescens (Makino) Nakai), 벚나무( P. jamasakura Siebold ex Koidz.), 개벚나무( P. verecunda (Koidz.) Koehne), 만첩개벚나무( P. verecunda var. semiplena (Nakai) W. T. Lee), 처진개벚나무( P. verecunda var. pendula (Nakai) W. T. Lee) 등 7분류군이다. 소메이요시노벚나무는 가로수나 정원수로 식재하는 벚나무류 중 가장 높은 비중을 차지하는 재배종으로 암술대에 털이 있고 악통이 컵 모양이며, 악편에 거치가 있는 특징으로 식별된다( Fig. 2A– D). 자생종인 왕벚나무와 비교하여 아린에 털이 밀생하는 특징으로 구분할 수 있으나, 이를 제외한 외부형태는 매우 유사하다( Fig. 2E– I). 형태적으로 왕벚나무는 개체 간 변이 폭이 크게 나타나는 데 비해 소메이요시노벚나무는 개체 간 차이가 거의 없는데( Kim et al., 1998), 이는 소메이요시노벚나무가 하나의 클론형을 가지는 재배종이기 때문이다( Iketani et al., 2007). 양벚나무는 속칭 ‘체리’로 유통되는 유럽과 서아시아 원산의 재배종으로 악편이 뒤로 젖혀지고 가장자리가 밋밋한 특징으로 소메이요시노벚나무와 구분된다( Fig. 2J, K). 처진올벚나무와 올벚나무는 모두 항아리 모양의 악통을 가지고 있으나, 처진올벚나무는 가지가 아래로 처지고, 올벚나무는 가지가 처지지 않는 특징에 의하여 서로 구분된다( Fig. 2L– N). 산벚나무와 벚나무는 식물체 전체에 털이 없고 악편에 거치가 없는 특징을 공유하며, 산벚나무는 대부분 총화경 길이와 분지하는 두 번째 소화경까지 길이가 짧아서 산형꽃차례이고( Fig. 2O– R), 벚나무는 산방화서이다( Fig. 2Z– AB). 왜벚나무는 악편에 거치가 있고 소화경에 털이 없으며( Fig. 2S– V), 잔털벚나무는 소화경에 털이 있는 특징으로 구분된다( Fig. 2W– Y). 왜벚나무는 일본 특산종으로 알려져 있었으나 비교적 최근에 국내 분포가 확인되었으며, 제주도에서 관상용으로 도입된 개체가 퍼져나간 것으로 추정되고 있다( Kim et al., 2011). 개벚나무는 산벚나무나 벚나무와 비교하여 소화경에 털이 듬성듬성 있는 점에서 구분되며( Fig. 2AC– AD), 원변종과 비교하여 꽃이 겹꽃인 종류는 만첩개벚나무, 가지가 아래로 처지는 종류는 처진개벚나무로 각각 구분된다( Fig. 2AE– AH). 다음은 식재 벚나무류 동정을 위해 작성한 벚나무아속(subg. Cerasus) 검색표이다. 조사지역 내 식재 벚나무류 중에는 아래 검색표에 의해 식별되지 않는 재배품종들이 있었는데, 본 연구에서는 미동정 분류군으로 처리하였다.
A key of Prunus subg. Cerasus in Korea
1. Style pubescent. 2. Calyx tube cup-shaped. 3. Calyx lobe straight, margin minutely serrate. 4. Outer bud scale hair dense ················································································· P. ×yedoensis 소메이요시노벚나무 4. Outer bud scale hair sparse ··································································································· P. ×nudiflora 왕벚나무* 3. Calyx lobe revolute, margin entire ····································································································· P. avium 양벚나무 2. Calyx tube vase-shaped. 5. Branchlets pendulous ·············································································································· P. spachiana 처진올벚나무 5. Branchlets ascending ································································································ P. spachiana f. ascendens 올벚나무 1. Style glabrous. 6. Calyx margin minutely serrate ······································································································· P. speciosa 왜벚나무 6. Calyx margin entire. 7. Pedicel glabrous. 8. Inflorescences corymbose with peduncles ·········································································· P. jamasakura 벚나무 8. Inflorescences umbellate with short peduncles or nearly no peduncles ····························· P. sargentii 산벚나무 7. Pedicel pubescent. 9. Inflorescences corymbose with peduncles ············································ P. serrrulata var. pubescens 잔털벚나무 9. Inflorescences umbellate with short peduncles or nearly no peduncles. 10. Branchlet straight. 11. Flowers single ················································································ P. verecunda var. verecunda 개벚나무 11. Flowers double ······································································ P. verecunda var. semiplena 만첩개벚나무 10. Branchlet pendulous ······································································· P. verecunda var. pendula 처진개벚나무 * not found in the survey area
벚나무류 식재 현황
식재된 벚나무류는 총 5,866주가 확인되었고, 이 중 5,744주는 11분류군으로 동정되었다( Fig. 2, Table 1). 확인된 벚나무류 중 소메이요시노벚나무가 3,055주(52.1%)로 가장 많았고, 다음으로 잔털벚나무 1,260주(21.5%), 벚나무 1,192주(20.3%) 순으로 이들 3종이 전체 93.9%에 이르며, 행정구역에 따른 식재 비율이 거의 차이가 없었다. 그 외 100주 미만이 확인된 벚나무류는 올벚나무 97주(1.6%), 처진올벚나무 42주(0.7%), 만첩개벚나무 32주(0.5%) 등의 순으로 확인되었고, 복합적인 특징을 나타내거나 알려진 특성과 일치하지 않아서 식별이 어려운 개체들은 122주(2.1%)가 확인되었다( Fig. 3, Table 1). 조사지역인 성남시 분당구 분당중앙공원 및 인접 지역은 대부분 주거용 건물 및 상가가 밀집되어 있고, 주로 개인주택 및 아파트단지, 학교 등에 녹지공간이 조성되어 있으며, 이들 도시녹지에 벚나무류를 도입할 때 소메이요시노벚나무가 가장 선호되는 수종으로 나타났다. 반면에 개체가 100주 미만으로 확인된 벚나무류 중 올벚나무는 분당동과 서현동 경계의 양영디지털고등학교 일대에 가로수로 연속 식재되어 있었고, 장안타운한신, 라이프빌라 내에도 다수가 식재되어 있었다. 분당중앙공원의 경우에는 조경수 식재지 31.1%, 잔존산림 51.3%, 수공간, 포장지, 건폐지 등 기타지역 17.6%로 구성되어 있으며, 1990년대 초 공원 조성 당시 남동쪽의 불곡산으로 이어지는 녹지축을 연결하기 위해 기존지형 및 수림대 보존 등 기존 환경을 최대한 활용할 목적으로 자생수목을 이식하고, 공원 남동쪽 분당천변 산책로에는 산벚나무를 식재하도록 계획하였다( Lee, 1995; Kang and Lee, 1996). 하지만, 현장 조사 결과 공원 남동쪽과 분당천 북쪽 산책로를 따라 분포하는 벚나무류는 잔털벚나무와 벚나무가 대부분이었으며, 소메이요시노벚나무가 매우 드물게 확인되었고, 산벚나무는 확인되지 않았다( Figs. 3, 4). 식재유형은 편의상 하천변 산책로 및 자동차 도로 주변에 1–2열의 선형으로 조성된 가로 식재와 공원 녹지대 내에 면으로 조성된 경관 식재 두 가지로 크게 나눌 수 있으며, 가로 식재의 경우 소메이요시노벚나무가 대부분으로 다른 수종이 매우 적고, 경관 식재의 경우 다양한 수종이 분포하는 것을 확인할 수 있었으며, 수형이 아래로 처지는 처진올벚나무와 처진개벚나무는 군식보다는 포인트 식재되며, 겹벚나무와 양벚나무는 경관 식재로 주거지 주변에 식재된 경우가 많았다( Fig. 3). 가로에 식재된 소메이요시노벚나무와 함께 확인된 벚나무류 수종은 접목으로 묘목을 생산하는 점을 고려했을 때, 증식 시 접목에 실패하여 대목이 성장한 개체 또는 종간 잡종이 함께 식재되었을 가능성도 있다고 여겨진다.
벚나무류 식재 관리 필요성
본 연구에서 확인된 벚나무류 외래수종은 3종류(소메이요시노벚나무, 왜벚나무, 양벚나무)이다( Table 1). 일본 원산 교잡종인 소메이요시노벚나무는 조사지역 내 벚나무류의 52.1%로 자생 벚나무류보다 큰 비중을 차지하였고, 전국에 가로수와 조경수로 많이 식재된 ‘왕벚나무’가 이에 해당하는 것으로 추정된다(Korea Forest Service, 2020) ( Table 1). 일본 고유종인 왜벚나무는 소메이요시노벚나무의 부계로 알려져 있다( Katsuki and Iketani, 2016). 왜벚나무의 정확한 도입 시기는 파악되지 않으나 제주도에서 관상용으로 식재되었다가 야생화된 것으로 보이는 개체군이 국내 미기록 귀화종으로 보고되었고, 본 연구에서 22주(0.4%)가 확인되었다( Kim et al., 2011) ( Table 1). 조사지역에서 가로수로 식재된 소메이요시노벚나무와 함께 왜벚나무가 발견되는 것은 녹지 조성 당시에 두 종의 뚜렷한 구분 없이 ‘왕벚나무’로 간주하였기 때문으로 보인다( Fig. 3D). 유럽과 서아시아 원산의 양벚나무는 과실수로 도입된 종으로서 관상용으로도 식재하며, 조사지역에서 16주(0.2%)를 확인하였다( Table 1). 양벚나무는 조사 지역 내에서 특정지역에만 확인된 것으로 보아 녹지 조성 당시에 의도적으로 식재된 것으로 추정된다( Fig. 3D). 이외에도 조경용이나 체리 생산을 위해 국내에 도입된 재배 품종은 약 50종류에 이른다( Lee et al., 2016). 본 연구에서 자생 벚나무류 및 주요 도입종의 식별형질과 정확히 일치하지 않는 122주는 비의도적으로 식재된 재배품종이나 종간 잡종으로 판단된다. 벚나무류는 비교적 쉽게 종간 잡종을 형성하며, 이러한 특성을 이용해 인공잡종을 형성하여 품종 개량을 시도하기도 한다( Liu et al., 2007). 반면, 중앙유럽의 희귀식물인 P. fruticosa Pall. 야생 개체군과 도입 재배종인 P. cerasus L. 사이에서 빈번하게 발생하는 잡종 형성과 역교배 사례연구는 식재된 개체군으로부터 야생 개체군에 유전자 이입이 발생하여 희귀식물의 고유성을 보전하는 데 심각한 위협 요소로 작용할 수 있다는 것을 보여준다(Macková et al., 2018). 즉, 벚나무류 자생지 인근에서 근연 외래종이 재배나 식재될 경우에 자생 벚나무류의 고유성 보전에 위협 요소로 작용할 수 있기 때문에 지양해야 한다. 개체군 크기가 작은 왕벚나무는 소메이요시노벚나무와 모계(올벚나무)가 같으며, 소메이요시노벚나무의 부계인 왜벚나무가 이미 유입되어 있어서 잡종을 형성할 가능성이 크다. 따라서 고유종인 왕벚나무 자생지 인근에 식재된 벚나무류의 적극적인 관리와 유전자 이입에 대한 모니터링이 필요하다. 잡종화와 유전자 이입에 의한 고유성 상실은 종간에 국한된 것은 아니며, 동일한 종 내에서도 유전적으로 격리된 집단에서도 적용된다( Rhymer and Simberloff, 1996). 따라서 식재 수목의 관리에는 종 식별과 원산지 파악이 중요하다. 실생묘 생산이나 재배종의 대목으로 산벚나무와 벚나무를 이용하는 것이 알려져 있을 뿐( Kim, 2004), 실체에 대해서 명확한 정보를 확인하지 못하였다. 산벚나무 종자는 주변국으로부터 수입되고 있는 실정으로서 이에 대한 실태 파악과 원산지 관리가 필요하다.
Fig 1.
Map of investigated area for this study. 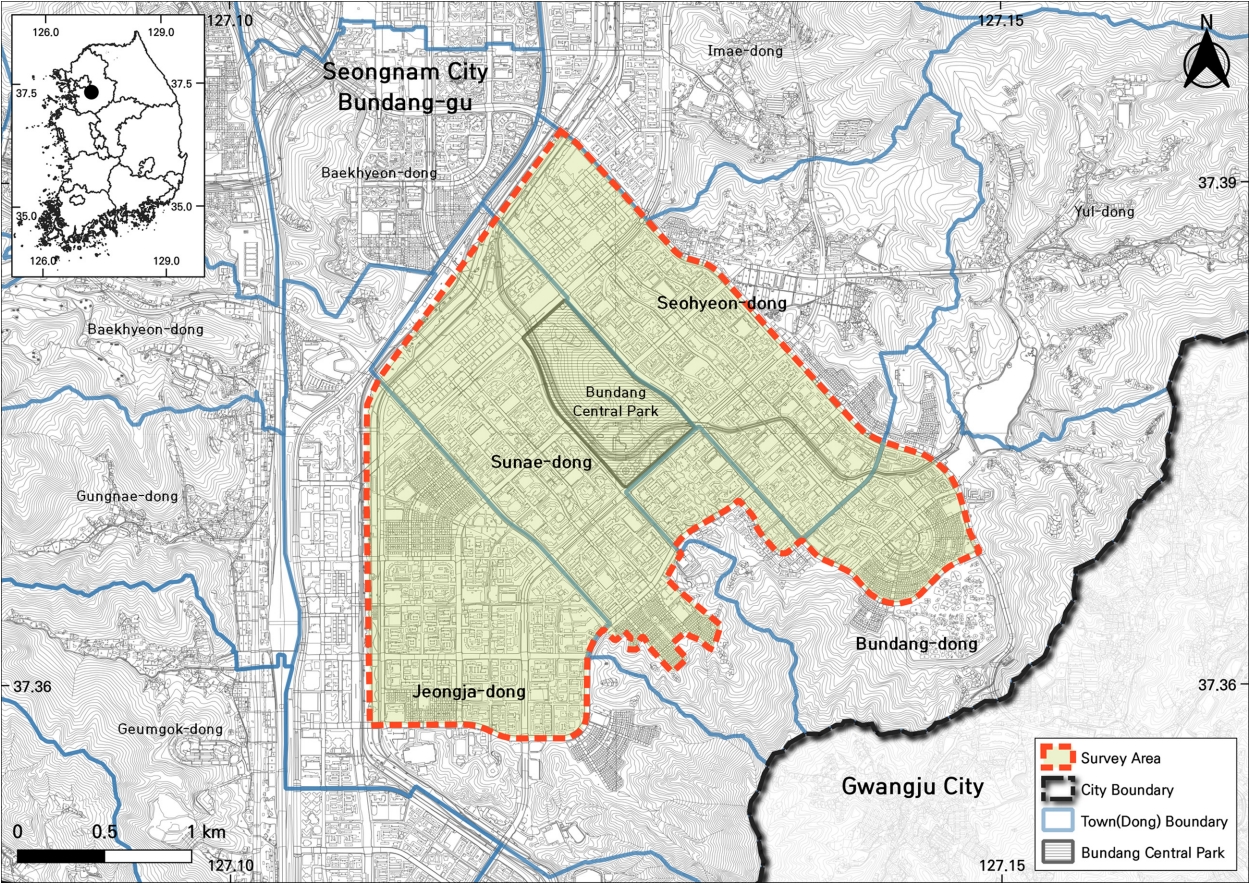
Fig 2.
Photographs of Prunus trees in Bundang Central Park and adjacent area. A–D. P. ×yedoensis. E–I. P. ×nudiflora (photographs taken on Jejudo Island). J, K. P. avium. L, M. P. spachiana. N. P. spachiana f. ascendens. O–R. P. sargentii. S–V. P. speciosa. W–Y. P. serrulata var. pubescens. Z–AB. P. jamasakura. AC, AD. P. verecunda. AE, AF. P. verecunda var. semiplena. AG, AH. P. verecunda var. pendula. A, F, P, T. Inflorescence. B, G, M, N. Calyx tube. C, H, R, V, Y, AB. Style (pubescent or glabrous). D, I. Bud scale. E, L, W, AG. Flowering branch. J, O, S, X, AA, AC, AD, AF, AH. Flowers. K. Calyx lobe (recurved). Q, U. Calyx lobe margin (entire or minutely serrate). Z. Inflorescence and young leaves. AE. Flower bud and 2 styles. 
Fig 3.
Distribution of Prunus trees in Bundang Central Park and adjacent area. A. P. ×yedoensis. B. P. serrulata var. pubescens. C. P. jamasakura. D. P. spachiana f. ascendens (black circle); P. spachiana (gray circle); P. verecunda var. semiplena (white circle); P. speciosa (black triangle); P. verecunda (gray triangle); P. avium (white triangle); P. sargentii (black square); P. verecunda var. pendula (gray square); Unidentified (white square). 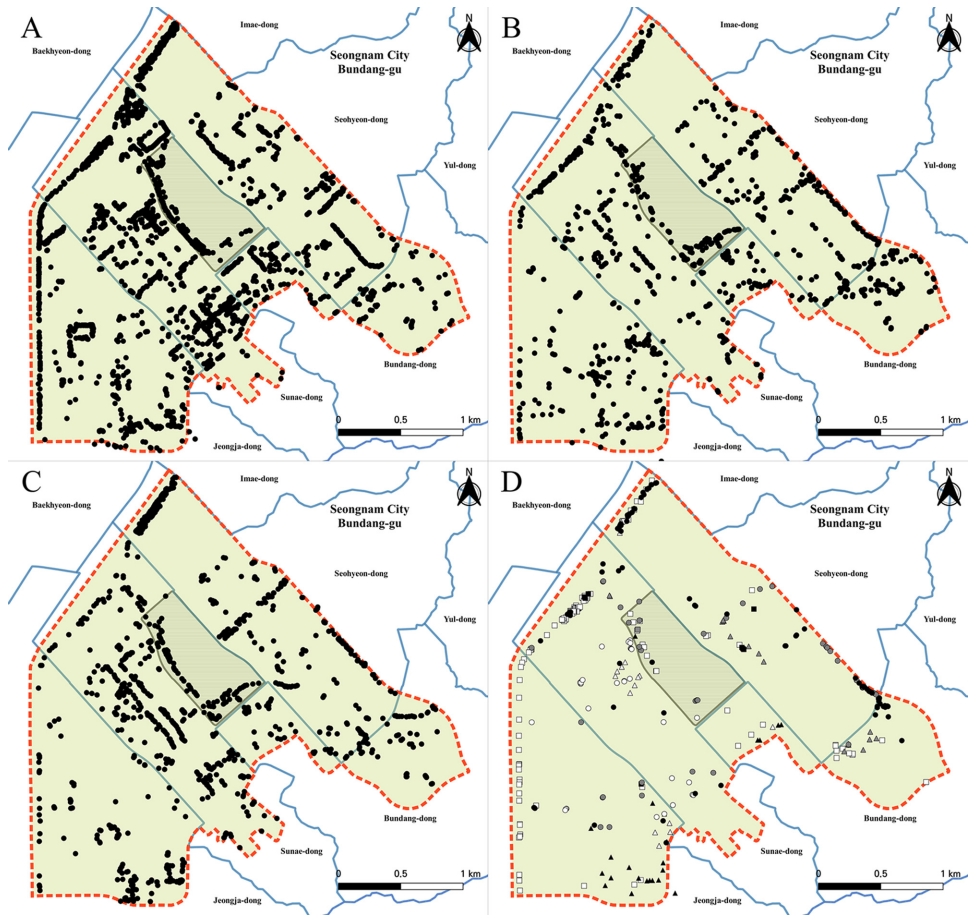
Table 1.
Current status of planted Prunus trees in Bundang-gu area.
|
Scientific name/ Korean name |
No. of trees |
Proportion (%) |
|
Bundang-dong |
Jeongja-dong |
Seohyeon-dong |
Sunae-dong |
Total |
|
Prunus ×yedoensis Matsum. 소메이요시노벚나무 |
338 |
589 |
780 |
1,348 |
3,055 |
52.1 |
|
P. serrulata Lindl. var. pubescens (Makino) Nakai 잔털벚나무 |
233 |
247 |
346 |
434 |
1,260 |
21.5 |
|
P. jamasakura Siebold ex Koidz. 벚나무 |
151 |
177 |
357 |
507 |
1,192 |
20.3 |
|
P. spachiana (Lavallée ex H. Otto) Kitam. f. ascendens (Makino) Kitam. 올벚나무 |
26 |
5 |
50 |
16 |
97 |
1.6 |
|
P. spachiana (Lavallée ex H. Otto) Kitam. 처진올벚나무 |
1 |
7 |
20 |
14 |
42 |
0.7 |
|
P. verecunda (Koidz.) Koehne var. semiplena (Nakai) W. T. Lee 만첩개벚나무 |
- |
6 |
- |
26 |
32 |
0.5 |
|
P. speciosa (Koidz.) Nakai 왜벚나무 |
4 |
17 |
- |
1 |
22 |
0.4 |
|
P. verecunda (Koidz.) Koehne 개벚나무 |
7 |
^— |
8 |
2 |
17 |
0.3 |
|
P. avium (L.) L. 양벚나무 |
1 |
5 |
2 |
8 |
16 |
0.2 |
|
P. sargentii Rehder 산벚나무 |
- |
1 |
6 |
3 |
10 |
0.2 |
|
P. verecunda (Koidz.) Koehne var. pendula (Nakai) W. T. Lee 처진개벚나무 |
- |
- |
- |
1 |
1 |
0.1 |
|
Unidentified |
13 |
31 |
20 |
58 |
122 |
2.1 |
|
Total |
774 |
1,085 |
1,589 |
2,418 |
5,866 |
100.0 |
Literature Cited
Baek, S.. Choi, K.. Kim, B.. Yu, J.. Cho, A.. Jang, H.. Kim, C.. Kim, J.. Chang, K.S.. Kim, H. and Mun, H.. 2018. Draft genome sequence of wild Prunus yedoensis reveals massive inter-specific hybridization between sympatric flowering cherries. Genome Biology 19: 127.   Chang, C.-S.. Choi, H. and Chang, S.. 2004. Reconsideration of Prunus sargentii complex in Korea: with respect to P. sargentii and P. takesimensis. Korean Journal of Plant Taxonomy 34: 221-244.  Cho, A.. Baek. G.-B. Kim, S.. Shin, H.. Kim, S.. Choi, K.. Kang, Y.. Yu, J.. Kim, H. and Mun, H.. 2017. Genomic clues to the parental origin of the wild flowering cherry Prunus yedoensis var. nudiflora (Rosaceae). Plant Biotechnology Reports 11: 449-459. Cho, M.-S. and Kim., C.. 2019. Multiple lines of evidence for independent origin of wild and cultivated flowering cherry ( Prunus yedoensis). Frontiers in Plant Science 10: 1555. Cho, M.-S.. Kim, S.. Kim, H. and Kim, C.. 2016. Comparative phylogenetic relationship between wild and cultivated Prunus yedoensis Matsum. (Rosaceae) with regard to Taquet's collection. Korean Journal of Plant Taxonomy 46: 247-255. Chung, G.Y.. Chang, K.S.. Chung, M.. Choi, H.J.. Paik, K. and Hyun, O.. 2017. A checklist of endemic plants on the Korean Peninsula. Korean Journal of Plant Taxonomy 47: 264-288 (in Korean). Chung, T.H.. 1957. Korean Flora I: Woody Plants. Sinjisa, Seoul. 589 (in Korean).
Chung, T.H.. 1965. llustrated Encyclopedia of Fauna and Flora of Korea, Vol. 5. Tracheophyta. The Ministry of Education, Seoul. 1824 (in Korean).
Chung, T.-H.. Do, S.. Lee, B. and Lee, J.. 1937. Botanical Names of Joseon. Society of History of Joseon, Seoul. 222 (in Korean).
Cultural HeritageAdministration. 2022. Korea National Cultural Heritage Portal. Retrieved Jan. 28, 2022, available from https://www.heritage.go.kr. Do, B.S.. Sim, H.J. and Im, R.J.. 1958. Illustrated Plant Book of Choseon (3). Institute of Science, Pyeongyang. 169 (in Korean).
Iketani, H.. Ohta, S.. Kawahara, T.. Katsuki, T.. Mase, N.. Sato, Y. and Yamamoto, T.. 2007. Analyses of clonal status in ‘Someiyoshino’ and confirmation of genealogical record in other cultivars of Prunus ×yedoensis by microsatellite markers. Breeding Science 57: 1-6. Ishidoya, T. and Chung., T.. 1923. The Identification Keys of Korean Forest Trees. The Government-General of Korea, Seoul. 129 (in Japanese).
Jin, E.-J.. Yoon, H.. Sung, H.. Bae, J.. Hyun, J.. Jung, Y. and Seo, O.. 2020. Comparison of flower morphological characteristics and palynological characteristics of native Prunus spp. in South Korea. Journal of Agriculture and Life Science 54: 11-20. Kalkman, C.. 2004. Rosaceae. The Families and Genera of Vascular Plants, Vol. 6. Flowering Plants, Dicotyledons. Kubitzki, K (ed.), Springer, Berlin. 343-374.
Kang, H.K. and Lee., K.J.. 1996. Establishment of ecological space for the connection of green corridor: a case study of ecobridge in Pundang, Songnam City. Korean Journal of Environment and Ecology 10: 24-38.
Katsuki, T. and Iketani., H.. 2016. Nomenclature of Tokyo cherry ( Cerasus ×yedoensis ‘Somei-yoshino’, Rosaceae) and allied interspecific hybrids based on recent advances in population genetics. Taxon 65: 1415-1419. Kim, C.S.. Lee, K.Y.. Moon, M.O.. Hyun, H.J.. Ihm, B.S. and Kim, M.H.. 1998. Natural habitat of Prunus yedoensis Matsumura and its morphological variation. Korean Journal Plant Taxonomy 28: 117-137. Kim, C.-S.. Moon, O.. Cheong, E.J. and Byun, G.O.. 2005. Evaluation of leaf morphology for distinguishing Prunus (Rosaceae) from Jeju, Korea. Korean Journal of Plant Taxonomy 35: 81-98. Kim, C.-S.. Moon, M.-O. and Kim, S.-Y.. 2011. A newly recorded naturalized species in Korea: Prunus speciosa (Koidz.) Nakai (Rosaceae, Prunoideae). Korean Journal of Plant Taxonomy 41: 253-258. Kim, S.I.. 2004. New landscaping tree: kinds of cherry tree and characteristics of growth. Landscaping Tree 79: 18-23 (in Korean).
Koehne, E.. 1912. . Prunus yedoënsis Matsum. var. nudiflora, nov. var. Repertorium Specierum Novarum Regni Vegetabilis. 10: 507 (in German).
Koidzumi, G.. 1932. Prunus yedoensis Matsum. is a native of Quelpaert. Acta Phytotaxonomica et Geobotanica 1: 177-179.
Korea Forest Service. 2020. National roadside tree furtherance achievements of 2019. Retrieved Jan. 14, 2022, available from https://www.forest.go.kr. Korea Forest Service. 2021. Forestry production survey of 2020. Retrieved Jan. 14, 2022, available from https://kfss.forest.go.kr. Korea National Aboretum. 2022. Checklist of vascular plants in Korea. Retrieved Jan. 28, 2022, available from http://nature.go.kr. Kuitert, W. and Peterse., A.H.. 1999. Japanese Flowering Cherries. Timber Press, Portland, OR. 395 pp.
Lee, C.R. and Ahn., H.S.. 1963. Nomina Plantarum Koreanum. Beomhaksa Publ. Co, Seoul, 525 pp. 525 (in Korean).
Lee, J.H.. Lee, S.Y.. Lee, K.C.. Kim, H.J.. Choi, H.. Lee, H.J.. Choi, K. and Chang, K.S.. 2016. Standard Checklist of Cultivated Plants in Korea. Sumeungil, Seoul. 510 pp.
Lee, J.-Y.. Park, S.. Kim, R.. D.-Y, . Kim, . Noh, S. and Lee, E.. 2011. Analysis of landscape planting in Gangwon-do. Journal of the Korean Institute of Landscape Architecture 39: 113-126. Lee, K.J.. 1995. Problems of provision for green space in new town: Bundang New Town. Review of Architecture and Building Science 39: 36-38 (in Korean).
Lee, S.T. and Kim., C.S.. 2007. Prunus L.. The Genera of Vascular Plants of Korea. Park, C.-W. (ed.), Academy Publishing Co, Seoul. 568-573.
Linnaeus, C.. 1753. Species Plantarum, Vol. 1.. Laurentius Salvius, Stockholm. 560 pp.
Liu, W.. Chen, X.. Liu, G.. Liang, Q.. He, T. and Feng, J.. 2007. Interspecific hybridization of Prunus persica with P. armeniaca and P. salicina using embryo rescue. Plant Cell Tissue and Organ Culture 88: 289-299. ;, L.. Vít, P. and Urfus, T.. 2018. Crop-to-wild hybridization in cherries: empirical evidence from Prunus fruticosa. Evolutionary Applications 11: 1748-1759. Matsumura, J.. 1901. Cerasi Japanicae duae species novae. The Botanical Magazine Tokyo 15: 99-101.
Milim Botanic Garden. 2022. List of commercial ornamental trees. Retrieved Jan. 28, 2022, available from http://milimbg.co.kr/. (in Korean). National Institute of Biological Resources. 2021. National list of species of Korea, 2020. Retrieved Jan. 28, 2022, available from http://kbr.go.kr. (in Korean). Northeastern Asia Biodiversity Institute. 2022. List of Korean vascular plants. Retrieved Feb. 19, 2022, available from http://koreanplant.info. (in Korean). Park, H.-S.. Lee, H.. Ahn, Y. and Kim, E.. 2000. Morphological and phenological comparisons of new Prunus species: a study on the flower, flowering time, and grafting efficiency. Journal of the Korean Institute of Landscape Architecture 28: 48-53.
Park, M.K.. 1949. An Enumeration of Korean Plants. Ministry of Education, Seoul, 516 pp. 516 (in Korean).
Rehder, A.. 1940. Manual of Cultivated Trees and Shrubs Hardy in North America Exclusive of the Subtropical and Warmer Temperate Regions. MacMillan, New York. 996 pp.
Rhymer, J.M. and Simberloff., D.. 1996. Extinction by hybridization and introgression. Annual Review of Ecology and Systematics 27: 83-109. Seongnam City. 2020. The Seongnam statistical yearbook of 2019. Retrieved Jan. 14, 2022, available from https://www.seongnam.go.kr/. (in Korean). Shi, S.. Li, J.. Sun, J.. Yu, J. and Zhou, S.. 2013. Phylogeny and classification of Prunus sensu lato (Rosaceae). Journal of Integrative Plant Biology 55: 1069-1079.
|
|











